
Pár hónapja írtam egy kutatásról, ahol a ma élő masszív bioinformatikai tour-de-force keretében a ma élő emlősök 99%-ból származó DNS mintákat összehasonlítva igyekeztek az emlősök kialakulásának történetét rekonstruálni.
Az eljárás alapjául az a megfigyelés szolgál, hogy az örökítőanyag változásai, vagyis a mutációk többé-kevésbe állandó ütemben rögzülnek, így ha sikerül rájönnünk mennyi időt jelent a molekuláris óra egy-egy "kettyenése", a DNS különbségek alapján megsaccolhatjuk, hogy mikor is élt a vizsgált fajok közös őse.
Ennek a korábbi tanulmánynak az volt a fő következtetése, hogy a főbb emlőscsoportok (kloakások, erszényesek és méhlepényesek) kialakulása bőven a Kréta és a Harmadkor (K/T) határán (~62 millió éve) bekövetkezett, hirtelen dinoszaurusz-kihalást megelőzően történt. Sőt, még az ősi méhlepényesek is megjelentek 150 millió éve, bár a mai fajokat kialakító radiációhoz még közel 100 millió évet kellett várni. A cikk így keményen szembement azzal a nézettel (amelyet főleg a fosszilis leletanyagból dolgozó paleontológusok favorizálnak), amely szerint az emlősök, de különösen a méhlepényesek elterjedése az említett K/T határára tehető, amikor a dinók kipusztulása életterek tucatjait tette elérhetővé az addig viszonylag pálya szélére szorított emlősök előtt.
Nem kellett sokat várni a viszontválaszra: a múlt heti Nature-ben jelent meg egy olyan cikk, amely szintén a lehető legteljesebb emlős törzsfát igyekszik elkészíteni - de DNS vizsgálatok helyett, az ismert Krétabeli emlős maradványok tüzetes elemzésével. Látható a fenti képen, hogy a két eljárás alapvetően nagyon hasonló ágazatú fát hoz létre (ami lényeges), egy fontos különbséggel: a fosszilis anyag szerint a méhlepényesek elterjedése mégiscsak a K/T határon történt és nem 40 millió évvel azt megelőzően.
Kinek van igaza és miért ez a nagy különbség? Az előbbi kérdésre lehetetlen válaszolni, csak feltételezni tudjuk, hogy valahol a kettő között keresendő a valódi dátum, de nem tudjuk melyikhez van közelebb. A különbséget pedig (feltehetőleg) a két módszer kisebb-nagyobb pontatlanságai okozzák.
A DNS vizsgálatok rendkívül informatívak, különösen nagy fajszám és sok vizsgált gén esetén, de ha ilyen időbeli következtetéseket akarunk levonni belőlük, akkor bizony pontosan kell tudnunk mennyi időt rejt egy-egy mutáció. Ez pedig azért nem mindig triviális, mert egy-egy csoport gyors radiációjakor gyorsabban fixálódhatnak a mutációk, mint egyébként, amit ha nem veszünk figyelembe, csalóka képet kaphatunk a rokoni kapcsolatok régiségéről. Ráadásul, ha egy bázis a DNS-ben A-ról C-re változik, majd néhány millió év múlva visszalakul A-ra, akkor az úgy is tűnhet, hogy nem volt semmilyen változás, ami szintén becslési hibát okozhat. Amint jobban megismerjük az efajta problémaforrásokat, megfelelő statisztikai módszerekkel tudunk majd korrigálni, de nem megy az egyik pillanatról a másikra.
De persze a fosszilis anyag vizsgálata sem maga a Kánaán: egy-egy csoportra jellemző jegyek alapján (pl. fogazat vagy belsőfül alakja) tudni fogjuk, hogy melyik az az időpont, amikor már biztos létezett az adott csoport, de nem lesz egyszerű megmondani, hogy mikor vált el a rokoncsoportoktól, amelyekkel való közös ősükben nem volt jelen a kulcsjegy. Ehhez még hozzáadva, hogy a fosszilizáció nem egy triviális folyamat, így elképzelhető, hogy bizonyos átmeneti formákat még nem leltek meg (és evolúciós időléptékben rövid ideig létezett fajoknál lehet, hogy nem is fognak), látható, hogy a klasszikus (paleo)taxonómusok dolga sem könnyű.
Szóval nem egyszerű ez. Nézeteltérés van, de ez egy nemes vita, amelynek a végén mindenki nyer majd: hiszen mindkét területnek alkalma lesz csiszolni és pontosítani saját eljárásait.
Cifelli, RL, Gordon, CL (2007) Re-crowning mammals. Nature 447: 918-920.
Wible, JR, Rougier, GW, Novacek, MJ, Asher, RJ (2007) Cretaceous eutherians and Laurasian origin for placental mammals near the K/T boundary. Nature 447: 1003-1006.

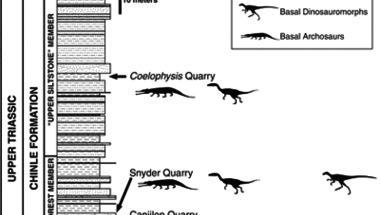
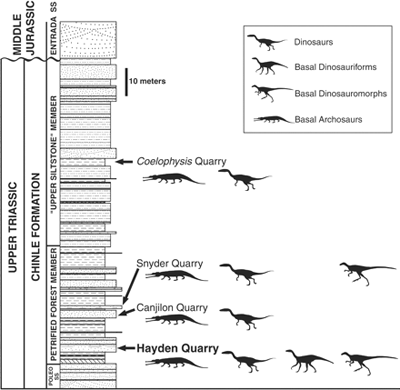 A dinoszauruszok a késői Triász folyamán jelentek meg (uszkve 200-230 millió éve) és ezt követően terjedtek el világszerte, fokozatosan kiszorítva más, primitívebb archoszaurusz csoportokat.
A dinoszauruszok a késői Triász folyamán jelentek meg (uszkve 200-230 millió éve) és ezt követően terjedtek el világszerte, fokozatosan kiszorítva más, primitívebb archoszaurusz csoportokat.
 Akik járatosak a témában, biztos felfigyeltek rá, hogy mikor a múlt héten az emberi sokféleség eredetét (pontosabban amit erről ma tudunk/gondolunk)
Akik járatosak a témában, biztos felfigyeltek rá, hogy mikor a múlt héten az emberi sokféleség eredetét (pontosabban amit erről ma tudunk/gondolunk) 
 Olvasom
Olvasom 
 Már régóta foglalkoztatja a történészeket, hogy vajon létezett-e, létezhetett-e kapcsolat a dél-amerikai kontinens és a polinéz szigetvilág lakói között, a Kolumbusz előtti időkben. Számos elmélet és cáfolat született eddig ezügyben (a leghíresebb talán Thor Heyerdahl
Már régóta foglalkoztatja a történészeket, hogy vajon létezett-e, létezhetett-e kapcsolat a dél-amerikai kontinens és a polinéz szigetvilág lakói között, a Kolumbusz előtti időkben. Számos elmélet és cáfolat született eddig ezügyben (a leghíresebb talán Thor Heyerdahl 


 Szűk egy éve
Szűk egy éve 
 Ha már néhány napja egy ismert ökológiai szabályt igazoló megfigyeléről
Ha már néhány napja egy ismert ökológiai szabályt igazoló megfigyeléről 
 Ha véginézünk a
Ha véginézünk a 
 A szigetek élővilága mindig különleges, gondoljunk csak Új Zéland egészen egyedi madár faunájára vagy Galapagos pintyeire és
A szigetek élővilága mindig különleges, gondoljunk csak Új Zéland egészen egyedi madár faunájára vagy Galapagos pintyeire és 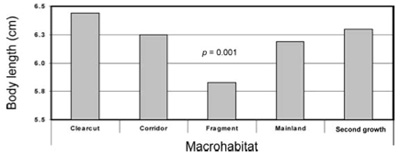 A Washington államban végzett kutatás végén a kutatók, több rágcsáló- (egér- ill. pocok-) fajt vizsgálva arra jutottak, hogy azok mérete az utóbbi mintegy három évtized alatt (ennyi a max. kora a vizsgált élőhely-töredékeknek) szignifikánsan csökkent más élőhelyen leledző rokonaikhoz képest. Mivel az egerek amúgy is igen sikeresek
A Washington államban végzett kutatás végén a kutatók, több rágcsáló- (egér- ill. pocok-) fajt vizsgálva arra jutottak, hogy azok mérete az utóbbi mintegy három évtized alatt (ennyi a max. kora a vizsgált élőhely-töredékeknek) szignifikánsan csökkent más élőhelyen leledző rokonaikhoz képest. Mivel az egerek amúgy is igen sikeresek 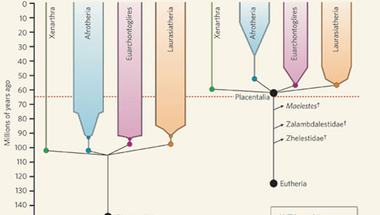
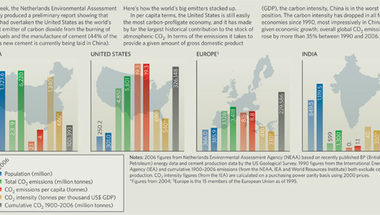
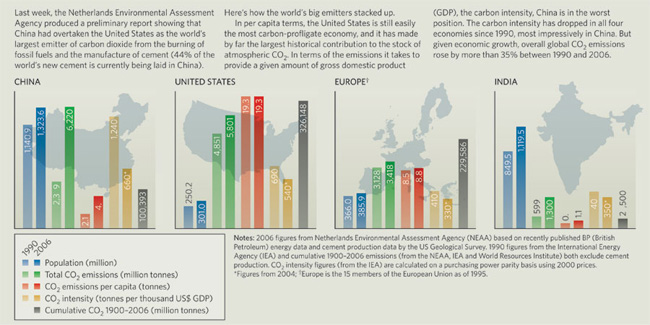

 Az emberi beszédnek számos genetikai komponense van: a légcső beszédhez szükséges anatómiai jellegzetességeinek fejlődése (akárcsak bármely más szervünk) egyértelműen számos gén interakciójától függ, és nincs ez nagyon másképpen az agy beszédképzéshez ill. -értéshez való "drótozásával" sem.
Az emberi beszédnek számos genetikai komponense van: a légcső beszédhez szükséges anatómiai jellegzetességeinek fejlődése (akárcsak bármely más szervünk) egyértelműen számos gén interakciójától függ, és nincs ez nagyon másképpen az agy beszédképzéshez ill. -értéshez való "drótozásával" sem.