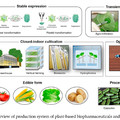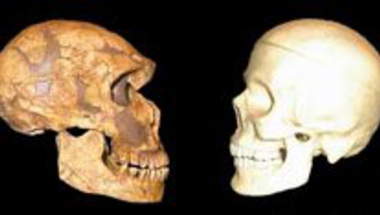
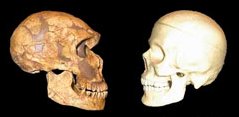
 Az emberiség bölcsője Afrika, méghozzá (a jelenleg legelfogadottabb elmélet szerint) többszörösen is. A kétlábra emelkedő Homo genus először úgy 1 millió éve kolonizálta az eurázsiai kontinenst a Homo erectus formájában, majd ezt újabb hullámok követték. Először fél millió éve a Homo heidelbergensis egy csoportja kelt útra, majd úgy 75-100,000 éve a modern ember őse, a Homo sapiens.
Az emberiség bölcsője Afrika, méghozzá (a jelenleg legelfogadottabb elmélet szerint) többszörösen is. A kétlábra emelkedő Homo genus először úgy 1 millió éve kolonizálta az eurázsiai kontinenst a Homo erectus formájában, majd ezt újabb hullámok követték. Először fél millió éve a Homo heidelbergensis egy csoportja kelt útra, majd úgy 75-100,000 éve a modern ember őse, a Homo sapiens.
Az afrikai szavannákról előbb a Közel Keletre vándorló (anatómiailag) modern emberek egy hosszabbacska ázsiai kitérő után érkeztek meg úgy 41,000 éve Európába. Itt pedig távoli rokonokkal futottak össze: az Afrikából korábban kivándorló H. heidelbergensis populációkból kifejlődő neandervölgyi emberekkel, azaz Homo neanderthalis-szal. Hogy pontosan mi következett ezután, az bősz találgatások és vita tárgya, mind a szak-, mind a bulvárirodalomban, mindenesetre tény, hogy 13,000 évvel később már csak az ibériai félszigeten létezett néhány neandervölgyi enklávé és azok sem sokáig. Sokak szerint a modern emberek brutálisan lemészárolták a neandervölgyieket, mások szerint (és ha engem kérdeznek ez a hihetőbb) a jobban alkalmazkodó H. sapiens szép csendben (és viszonylag békésen) szorította ki unokatestvérét.
Az egyik legérdekesebb és jócskán vitatott kérdés, hogy mennyire voltak csapodárok a két Homo populáció tagjai, vagyis volt-e keveredés közöttük.
Pusztán morfológiai jelek alapján valószínűleg ezt lehetetlen eldönteni, éppen ezért kapóra jön egy jó kis genom project. Nameg, arra is alkalmas lehet a neandervölgyi ember genomja, hogy még pontosabb képünk legyen arról, hogy melyek lehettek azok a genetikai változások, amelyek az emberek őseit végül elválasztották a csimpánzok őseitől (az sem ment éppen ripsz-ropsz), illetve mely változások tették igazán különlegessé a H. sapiens vonalát. (Csak arra nem kaphatunk pontos választ, hogy mely változások tették a H. neanderthalis vonalat különlegessé, de ezt lásd később.)
A genom projecthez egy horvát (vindijai) fosszíliát használtak, és ebből izolált mindkét kutató csoport DNS mintát. A neheze azonban ezután következett: a szolid 38,000 földben töltött év a csontot mindenféle baktériumok, gombák és más mikroszkópikus élőlények kolonizálták be. Ráadásul a DNS egy törékeny jószág, így az egykori genom már apró 50-100 bázispár hosszú darabokra hullott szét. Így az izolált össz DNS apró darabjait megszekvenálva el kellett dönteni, hogy az érdekes-e a kutatás szempontjából, vagy csak valami szennyeződés. Végül a DNS kb. 6,2%-a bizonyult igazán érdekesnek, mert ez hasonlított legjobban a két referencia genomhoz, az emberéhez és a csimpánzéhoz. Ezeket egyébként egyfajta vázként használták, hogy megtalálják az apró neandervölgyi DNS darabok genomon belüli helyét (hiszen ez a genom is nagyon hasonló kell legyen az egymsához is hasonlító referencia genomokhoz).
A szekvenáláshoz két csapat látott hozzá, az egyik egy hagyományos módszert alkalmazva klónozta az apró darabokat mesterséges bakteriális vektorokba és ezt követően szekvenálta meg (ők 65,250 bázispárig jutottak így), a másik társaság azonban igazi technológiai tour de force mutatvánnyal rukkolt elő. Ennek során egy ötletes eljárással apró golyókhoz kötötték egyenként a DNS darabokat és a golyók felszínen sokszorosították, majd szekvenálták őket. Mivel az eljárás teljes egészében automatizálható, lényegesen impozánsabb 1 millió bázispárt sikerült így meghatározni (becsléseik szerint az egész genomot sikerül 2 éven belül lefedniük ezzel a módszerrel).
Az eredmények egy része nem meglepő, szinte elvárt volt: ilyen pl., hogy a mi genomunk és alacsony homlokú, de nagy koponyájú rokonainké 99.5%-ban azonos (a csimpánz esetében ez a szám 95% körül van). Ők is, akárcsak jómagunk viszonylag kisméretű (pár ezer fős) kiindulási populációból származnak. Amire azonban még mindig nincs egyértelmű válasz az az esetleges flörtölések kérdése. Ennek eldöntésére az elmúlt években több neandervölgyi fosszília mitokondriális DNS-ét szekvenálták meg. A mitokondriumok sejtjeink apró energiagyárai, és jellemző tulajdonságuk, hogy apró saját genetkai állományuk van, és csak anyai ágon öröklődnek. Nos, az eddig megszekvenált 12 fosszilis H. neanderthalis mitokondrium DNS (mtDNS) genom, jellemző helyeken különbözött az emberi genomtól, de egymáshoz ugyanott meg nagyon hasonlítottak. Ez a megfigyelés csökkentette a keveredés esélyét, ha nem is zárta ki teljesen. Nem zárhatta ki, ugyanis pont az öröklődési mechanizmus amiatt, az esetleges neadnervölgyi mtDNS kihígulhatott, hiszen ha valahol megszakad a lány leszármazás ág ez a genetikai anyag nem adódik át, vagyis elvesződik. A vita végleges lezárásához így szükség van a sejtmagban meghúzódó és "mendelien" öröklődő genetikai állomány vizsgálatához.
Akad azonban egy másik nehezítő körülmény: a DNS nemcsak fragmentálódik az idők folyamán a csontokban, de egyes bázispárok előszeretettel elkezdenek mutálódni is (a legagykoribb a C->T, ill. G->A átalakulás). Ez igaz az mtDNS-re is, de mivel sejtenként több ezer mitokondrium van, az egyes hibák könnyen kiszűrhetők. A sejtmagban meghúzódó DNS esetében azonban nincs ilyen luxus. Éppen ezért azok a bázispárok, ahol a neandervölgyi DNS mind a homológ csimpánz, mind a homológ sapiens DNS szakaszoktól különbözik, nehezen értékelhetőek, hiszen nem tudhatjuk, hogy a különbség a lebomlásnak tudható be, vagy valóban létező eltérés okozta. Így aztán marad annak vizsgálata, hogy adott helyeken a csimpánz-, vagy a modern emberi genommal egyezik meg a neandervölgyi DNS szekvencia.
Ez azonban nem lebecsülendő, hiszen rengeteg információt hordoz. Különösen azok a helyek izgalmasak, ahol az emberi genomban is polimorfizmusok fordulnak elő, vagyis két ma élő ember között is jó eséllyel különbözik egy-egy bázispárban az adott DNS szakasz. Ezeket az apró eltéréseket nevezzük SNP-nek (single nucleotide polymorphism), és ezek vizsgálatára, vagyis az emberi diverzitás feltérképezésre indították anno a HapMap projectet. Egy-egy SNP-nek létezik "ősi" változata, amikor a változékony bázispár kvázi megegyezik a csimpánzban előfordulóval, máskor azonban különbözik attól, ez az ún. "számaztatott" formája. Hogy a származtatott forma mennyire gyakori az egész emberiségen belül, az sok mindentől függ, de legfőképpen attól, hogy milyen hamar jelent meg a csimpánz-ember szétválás után. Minnél hamarabb, annál elterjedtebb lesz. Jó, de hogy jön ez a dolog a neandervölgyiekkel való esetleges unga-bungához, kérdezheti a nyájas olvasó? Hát itt éppen úgy, hogy megvizsgálták, hány esetben fordul elő a származtatott forma a neandervölgyi genomban, mintegy ezer SNP esetében. A kapott 30% pedig több, mint amit egy egyszerű félmillió éves végleges szétválás esetében feltételezhetnénk. (Elsősorban az európaiakban gyakori SNP-kre kell koncentrálni, hiszen az esetleges keveredés velük történhetett, az afrikaiak azon anatómiailag modern emberek leszármazottai akik a legutolsó nagy kivándorlási hullmhoz nem csatlakoztak, így értelemszerűen az nekik fizikailag nem volt esélyük szerelmi légyottokat folytatni a neandervölgyiekkel.)
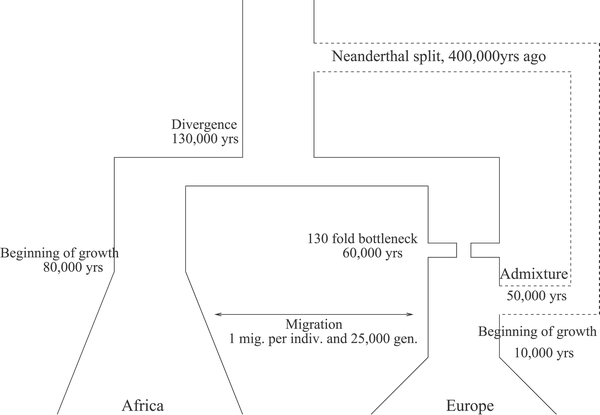
Az adatok alapján inkább tűnik valószínűnek, hogy mégis volt valami egymásba gabalyodás a két Homo faj egyedei között. És mivel a neandervölgyi X kromoszóma jobban különbözik, mint az autoszómái a mi genetikai állományunktól, elképzelhető, hogy a kavarodásért elsősorban, khmmm, a H. sapiens férfiak voltak a felelősek...
Egy másik, kicsit indirektebb bizonyíték a microcephalin génhez kapcsolódik. Ennek a génnek fontos szerepe van az agyméret szabályozásában, ugyanis ha valami miatt nem működik, az az agy drasztikus csökkenését okozza (bár érdekes módon szerkezetileg nem lesz benne sok változás). Talán nem túl meglepő módon a csimpánz-ember vonalak egykori szétválása után a gén erőteljes változásba kezdett az emberi vonalon, és ez a feltételezhető (egyik) oka annak, hogy az emberi agy lényegesen nagyobb, mint legközelebbi élő rokonunké.
Ugyanakkor az emberi populációkban sem totál egyforma a microcephalin gén, két alléja ismert: az egyik az ún. D haplotípus, a másik pedig a non-D haplotípus, amelyek szekvenciájukban egyetlen aminosavban különböznek. Eurázsiában és az Amerikai kontinensen egyaránt a D haplotípus az elterjedt, míg Afrikában a non-D. Emiatt, amikor az eredeti cikk megjelent néhány zseni igyekezett felhasználni beteges rasszista elméletének megtámogatására. Éppen ezért hangsúlyoznom kell, hogy jelenleg fogalmunk sincs mi a működésbeli különbség a két allél között, semmi alapunk azt hinni, hogy az egyiket hordozó egyén okosabb mint a másikkal rendelkező társa. Ugyanakkor kétségtelen, hogy a non-D haplotípusnak valamilyen szelekciós előnye van az említett területeken, másképp nem magyarázható az elterjedtsége (a teljes emberi népességben 30%-70% a non-D - D arány).
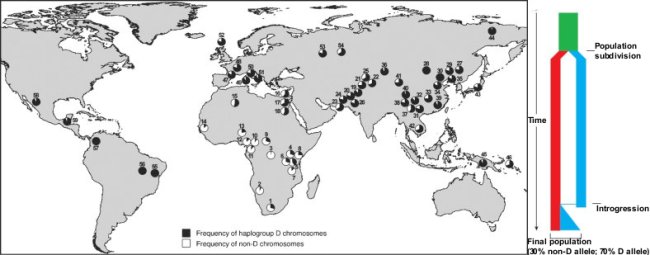
A D-haplotípus további érdekessége, hogy ha a microcephalin gén nem kódoló részeit is megvizsgáljuk az említett aminosav eltérést okozó mutáció környékén, számos további SNP-t fedezhetünk fel, amelyek specifikusak a D-haplotípusra nézve. Olyannyira, hogy ha egészen eltérő populációkban összehasonlítjuk az említett szakaszokat, azok alig különböznek (márpedig hosszú idő alatt a nem kódoló részekben viszonylagos szabadsággal jelenhetnek meg apró változások), ami arra utal, hogy ez a haplotípus mindössze ~37,000 éve jelent meg. A korábban elmondottak fényében ez már eleve gyanús lehet, de még semmit sem bizonyít :-). Az azonban fokozza a gyanút, hogy ha magát a teljes D-haplotípus specifikus DNS szakaszt (kódoló és nem kódoló részeket egyaránt) hasonlítjük össze a non-D megfelelőjével rengeteg különbséget lelhetünk, mondhatni meglepően sokat. Sokkal többet, mint amit az ősi és változékonyabb non-D haplotípuson belül találunk. Ez pedig csak úgy magyarázható, hogy a két allél eredetileg több mint félmillió éve különvált, és elszigetelt Homo populációkban fejlődött (az egyik, a non-D-hez kapcsolódó, az a modern emberősök csoportja, a másik pedig kérdőjel), majd sokkal később a populációk újból találkoztak és keveredtek. Ekkor egy introgressziónak nevezett jelenség során a microcephalin D allélja visszakerült a H. sapiens populációba. S mivel a már említett, egyelőre titokzatos szelekciós előnyt biztosította a szóbanforgó területeken, gyorsan rögzült és el is terjedt.
Persze nem árt kiemelni, hogy a microcephalin kicsit kakukktojás, a legtöbb gén esetében nem találkozunk hasonló kacifántos történettel. De éppen ettől válik ez a gén érdekessé.
 És hogy helyileg hol zajlott vajon a neandervölgyiek és modern emberek keveredése? Erre szekvencia adatunk kevés van, viszonyt számos olyan csontmaradvány került elő Európa szerte, amelyek két Homo faj jegyeinek érdekes keveredését mutatják. Legutóbb például (hogy egy mind időben, mind térben közeli példát hozzak) a romániai Gorzs megyében található Vénasszony barlangban (Pestera Muierii) bukkantak olyan koponyára, amelynek arckoponyája inkább modern emberre, agykoponyája pedig inkább neandervölgyire emlékeztet.
És hogy helyileg hol zajlott vajon a neandervölgyiek és modern emberek keveredése? Erre szekvencia adatunk kevés van, viszonyt számos olyan csontmaradvány került elő Európa szerte, amelyek két Homo faj jegyeinek érdekes keveredését mutatják. Legutóbb például (hogy egy mind időben, mind térben közeli példát hozzak) a romániai Gorzs megyében található Vénasszony barlangban (Pestera Muierii) bukkantak olyan koponyára, amelynek arckoponyája inkább modern emberre, agykoponyája pedig inkább neandervölgyire emlékeztet.
Persze, akárcsak a "hobbitok" esetében, a morfológiai jegyek értelmezése széles vitaalapot biztosít, ezért végleges választ arról, hogy a koponya egykori tulajdonosa mennyire volt neandervölgyi csak a DNS vizsgálatok adhatnak majd.
A neandervölgyi genom titkai még csak most kezdtek feltárulni és biztos, hogy a következő két évben hallani fogunk olyan génjeikről, amelyek nagyon fontosak a modern emberek számára, pl. a beszédkészség szempontjából nélkülözhetetlen FOXP2 génről. Vajon milyen volt ez távoli unokatestvéreinkben...?
Bővebben: nature:neanderthaldna és John Hawks blogja.
Green RE, Krause J, Ptak SE, Briggs AW, Ronan MT, Simons JF, Du L, Egholm M, Rothberg JM, Paunovic M, Pääbo S. (2006) Analysis of one million base pairs of Neanderthal DNA. Nature 444: 330-336.
Noonan JP, Coop G, Kudaravalli S, Smith D, Krause J, Alessi J, Chen F, Platt D, Pääbo S, Pritchard JK, Rubin EM. (2006) Sequencing and analysis of Neanderthal genomic DNA. Science 314: 1113-1118.
Plagnol V, Wall JD(2006) Possible ancestral structure in human populations. PLoS Genet 2(7): e105. DOI: 10.1371/journal.pgen.0020105
Evans PD, Gilbert SL, Mekel-Bobrov N, Vallender EJ, Anderson JR, Vaez-Azizi LM, Tishkoff SA, Hudson RR, Lahn BT. (2005)Microcephalin, a gene regulating brain size, continues to evolve adaptively in humans. Science 309: 1717-1720.
Evans PD, Mekel-Bobrov N, Vallender EJ, Hudson RR, Lahn BT. (2006) Evidence that the adaptive allele of the brain size gene microcephalin introgressed into Homo sapiens from an archaic Homo lineage. PNAS DOI: 10.1073/pnas.0606966103
Soficaru A, Dobos A, Trinkaus E. (2006) Early modern humans from the Pestera Muierii, Baia de Fier, Romania. PNAS 103(46): 17196-17201.