Ha véginézünk a jelenleg (genom)szekvenálás alatt álló állatok során, akkor az egyik első dolog ami feltűnhet, az a listát alkotó fajok színes, látszólag minden logikát nélkülöző kavalkádja. Vajon miért pont a tatu és mókus, a kacsacsőrű emlős és az ingola, a tenrek és a zsákállat került szekvenátorvégre? Hiszen annak látható a logikája és potenciális haszna, hogy az orvosi kutatások modellállatainak (egér, Drosophila, C. elegans, zebrahal), illetve az emberiség fontos haszonállatainak (disznó, szarvasmarha, ló) a genomját a kezünkben akarjuk tartani, de mi a haszna a fent felsoroltaknak? Ez már a "szekvenáljunk meg minden fajt" fázis lenne, ahol a kutatók azt vizsgálják, ami éppen a kezük ügyébe akad?
Ha véginézünk a jelenleg (genom)szekvenálás alatt álló állatok során, akkor az egyik első dolog ami feltűnhet, az a listát alkotó fajok színes, látszólag minden logikát nélkülöző kavalkádja. Vajon miért pont a tatu és mókus, a kacsacsőrű emlős és az ingola, a tenrek és a zsákállat került szekvenátorvégre? Hiszen annak látható a logikája és potenciális haszna, hogy az orvosi kutatások modellállatainak (egér, Drosophila, C. elegans, zebrahal), illetve az emberiség fontos haszonállatainak (disznó, szarvasmarha, ló) a genomját a kezünkben akarjuk tartani, de mi a haszna a fent felsoroltaknak? Ez már a "szekvenáljunk meg minden fajt" fázis lenne, ahol a kutatók azt vizsgálják, ami éppen a kezük ügyébe akad?
Nos, bár a távlati cél tényleg minden faj DNS-ének feltérképezése (sőt, minden faj több egyedének a vizsgálata, hiszen a fajokon belüli variancia gyakran uo. érdekes, mint a fajok közötti), a listára jelenleg korántsem véletlenszerűen lehet felkerülni. Annyira ugyanis még nincs szekvenáló-kapacitás fölösleg, hogy efajta luxust engedélyezhessenek maguknak a genetikusok; a mai törekvések fő célja az, hogy minnél jobban megértsük a már rendelkezésünkre álló genomokat, de különösen a saját magunként (ennyi önzőzés talán minden fajtól megengedhető ;-)). Ehhez pedig arra van szükség, hogy fokozatoan egyre távolabbi rokonaink genomját tárjuk fel, mert minden egyes ilyen lépéssel egy kicsit pontosabban látjuk, mikor, melyik más csoportokkal való közös ősben jelentek meg egyes jellegzetes DNS szekcenciák, azaz hogyan farigcsálta a természetes szelekció révén az evolúció a genetikai állományunkat.
S, hogy mennyire fontos, minnél több fajból meríteni, azt mi sem példázza jobban, mint az, ha felidézzük, hogy az első pár többsejtű-genom publikálása után, a kis mintaszám miatt, milyen hibás elméletek születtek.
Ha véginézünk a jelenleg (genom)szekvenálás alatt álló állatok során, akkor az egyik első dolog ami feltűnhet, az a listát alkotó fajok színes, látszólag minden logikát nélkülöző kavalkádja. Vajon miért pont a tatu és mókus, a kacsacsőrű emlős és az ingola, a tenrek és a zsákállat került szekvenátorvégre? Hiszen annak látható a logikája és potenciális haszna, hogy az orvosi kutatások modellállatainak (egér, Drosophila, C. elegans, zebrahal), illetve az emberiség fontos haszonállatainak (disznó, szarvasmarha, ló) a genomját a kezünkben akarjuk tartani, de mi a haszna a fent felsoroltaknak? Ez már a "szekvenáljunk meg minden fajt" fázis lenne, ahol a kutatók azt vizsgálják, ami éppen a kezük ügyébe akad?
Nos, bár a távlati cél tényleg minden faj DNS-ének feltérképezése (sőt, minden faj több egyedének a vizsgálata, hiszen a fajokon belüli variancia gyakran uo. érdekes, mint a fajok közötti), a listára jelenleg korántsem véletlenszerűen lehet felkerülni. Annyira ugyanis még nincs szekvenáló-kapacitás fölösleg, hogy efajta luxust engedélyezhessenek maguknak a genetikusok; a mai törekvések fő célja az, hogy minnél jobban megértsük a már rendelkezésünkre álló genomokat, de különösen a saját magunként (ennyi önzőzés talán minden fajtól megengedhető ;-)). Ehhez pedig arra van szükség, hogy fokozatoan egyre távolabbi rokonaink genomját tárjuk fel, mert minden egyes ilyen lépéssel egy kicsit pontosabban látjuk, mikor, melyik más csoportokkal való közös ősben jelentek meg egyes jellegzetes DNS szekcenciák, azaz hogyan farigcsálta a természetes szelekció révén az evolúció a genetikai állományunkat.
S, hogy mennyire fontos, minnél több fajból meríteni, azt mi sem példázza jobban, mint az, ha felidézzük, hogy az első pár többsejtű-genom publikálása után, a kis mintaszám miatt, milyen hibás elméletek születtek.
A C. elegans nevű fonalféreg genom (egyébként ez volt az első többsejtű genom) 1998-as, majd a muslica (Drosophila) és humán genomok 2000-es bejelentése után, elindult a vad összehasonlítgatás, miben is különbözünk mi, az "újszájúak" deli képviselője, ettől a két "ősszájú"-tól. A legnagyobb visszhangot az kapta, hogy mennyire meglepően hasonló számú (fehérje kódoló) gén fordul elő a három genetikai állományban, s az már csak a kevésbé populáris cikkekben került megemlítésre, hogy ezeknek a géneknek a szerkezete azonban gyakran különböző.
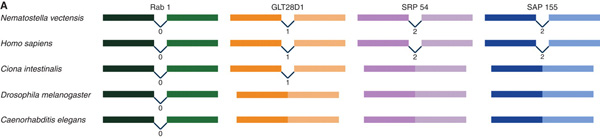
A legtöbb fehérje-kódoló gén DNS szekvenciájában a tankönyvi felosztás szerint három fő régiót különböztethetünk meg (megj.: az elmúlt években elég sokat változott a gén-definíció, az itt használt már egy kicsit inkább csak részhalmaza annak, amit ma alkalmaznak a biológusok, de itt a célnak megfelel): a gén átíródását szabályozó promotereket, a konkrét fehérjeszekvenciát rejtő exonok, és az utóbbiak között elhelyezkedő intronok. Az intronok, a DNS -> RNS átíródás után, egy speciális folyamat, a splicing keretében kivágódnak, így aztán a végtermék az exonoknak megfelelő szekvenciák összefűzéséből létrejövő mRNS lesz.
A Drosophila és C. elegans vs. humán genom összehasonlítások egyik feltűnő hozadéka volt, hogy az emberi genom rendre több intront tartalmazott, mint a másik kettő. A különbség okaira több elmélet is született, s az eredetét sokan annak tulajdonították, hogy a kétoldali-szimmetriájú állatok közös ősében (ún. Urbilateria) kevés intron volt, s ezek majd később, a gerincesek evolúciója során alakultak ki.
Az elmélet viszonylag rövid életűnek bizonyult, fokozatosan gyűltek az ellenérvek és végül a kegyelemdöfést bő másfél éve egy másik ősszájú, egy tengeri gyűrűsféreg, a Platynereis dumerilii genomjának vizsgálata adta meg. Ugyanis ebben a furcsa kis élőlényben, nemcsak, hogy szintén sok intron volt, de azok gyakorlatilag bázispárra megegyezően ott helyezkedtek el a DNS-ben, mint a mi intronjaink. Márpedig annak az esélye, hogy ezek egymástól függetlenül létrejöjjenek, elég jó közelítéssel nulla, azaz minden jel szerint már a mitikus Urbilateriában is ugyanott voltak jelen mint bennünk, csak a fonálférgek és a muslicák vonalában elvesztődtek.
Mi vezette félre akkor először a kutatókat, miért ennyire más ezek szerint a Drosophila és C. elegans genom? Utólag könnyű okosnak lenni, s retrospektíve már látható, miért kellett volna óvatosnak lenni az első genom-összevetések során: mindaz ami ezeket az állatokat ideális modell-organizmussá teszi (gyors fejlődés és generációs idő), egyszersmind nyomot hagy a genomjukon is. A gyors fejlődés általában különleges megoldásokat igényel a testtengelyek embrionális kialakítása során és a sejtosztódások közben. Pl. minnél kevesebb a genetikai anyag, annál gyorsabban (és kevesebb energia felhasználásáva) osztódik a sejt, így annál gyorsabban fejlődhet az embrió is. Vagyis az egyik lehetséges megoldás megszabadulni az olyan DNS szakaszoktól, amelyek nem létfontosságúak - és a hiányzó intronok éppen ebbe a kategóriába estek.
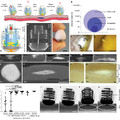
A Platynereis-es, intronok ősiségét igazoló tanulmány a héten újabb támogatást kapott, mégpedig úgy, hogy még egy lépést tettünk visszafelé a törzsfejlődés időskáláján és a "körkörös-" (Radiata) ill. "kétoldali" szimmetriájú (Bilateria) állatok közös ősének genetikai felépítését vizsgálandó, befejeződött az előbbi csoportba tartozó kis csalánozó, a Nematostella vectensis genomjának a szekvenálása.
A csalánozók (Cnidaria) kis virágállatok, hidrák és medúzák, nagyon egyszerű felépítésű élőlények, és bár kétségtelenül valódi szöveteik vannak, máig nem eldöntött, hogy ezek két vagy három csíralemezből jönnek ltre, vagyis van-e valódi mezodermájuk (nálunk gerinceseknél ebből a csíralemezből jönnek létre a csontok, az izmok, a vér, és a szaporódó- és kiválasztó készülék). Mégis, a jelek szerint a szervezeti egyszerűség ellenére, genomjuk közel annyira komplex mint a miénk, vagy a Platynereis-é.
Az eredmények nem voltak teljesen váratlanok, mert az első szekvenciák befutása után már sejteni lehetett, hogy azok a kis egyszerű virágállatok, nem is annyira egyszerűek (ui. a legtöbb fontos transzkripciós faktor- és jelátviteli útvonal-komponens génje ott figyelt a DNS-ükben), de azért a teljes genom is mindenképpen említést ér.
A legérdekesebb, bár a fentiek fényében már talán nem is meglepő, hogy az intronjaik ~80%-a ugyanott található, mint bennünk, emberekben. Vagyis, ezek szerint nemcsak az Urbilateria, de a szövetes állatok, röpke 700 millió éve élt őse is rendelkezett velük. Ennél váratlanabb volt, hogy a mintegy 18,000 fehérje kódoló génjük közül (csak az összehasonlítás véget, az emberi genomban a mai saccok szerint kb 20,000 - 23,000 van...) nagyon sok hasonló szomszédokkal van övezve, mint a mi genomunkban (ezt a jelenséget szinténiának nevezzük). A teljes genom (~450 millió bázispár) mintegy negyede áll ugráló gének maradékából (nálunk ez 50% körül van).
Szóval ezek a csalánozók igazán különlegesek, és még biztosan tartogatnak meglepetéseket. Például az egyik nagy vita, amiről korábban már szintén volt szó, hogy valóban körkörösen szimmetrikusak-e, vagy a látszat csalóka. Ez azért merült fel, mert molekulárisan számos gén kétoldali szimmetria alapján fejeződik bennük ki, azaz egyesek szerint az sem kizárt, hogy a látszólagos körkörösség csak másodlagos (derivált) tulajdonság és a csalánozók őse maga valójában kétoldalian szimmetrikus volt.
 S ha ez még mind nem lenne elég a csalánozó-különlegességekből, akkor, itt van ez a furcsa kis féregalakú lény a baloldali képen. Ez egy Buddenbrockia plumatellae, egy mikroszkópikus méretű kis parazita, ami a rejtélyes Myxozoa csoportba tartozik. A csoport taxonómiailag nem teljesen koherens, inkább csak gyűjtőhelye a nehezen beazonosítható, apró élősködőknek (sok helyen egysejtűként definiálják őket, pl. Wikipédia, ami nem felel meg a valóságnak).
S ha ez még mind nem lenne elég a csalánozó-különlegességekből, akkor, itt van ez a furcsa kis féregalakú lény a baloldali képen. Ez egy Buddenbrockia plumatellae, egy mikroszkópikus méretű kis parazita, ami a rejtélyes Myxozoa csoportba tartozik. A csoport taxonómiailag nem teljesen koherens, inkább csak gyűjtőhelye a nehezen beazonosítható, apró élősködőknek (sok helyen egysejtűként definiálják őket, pl. Wikipédia, ami nem felel meg a valóságnak).
Most, mintegy 50 db. fehérje-kódoló génjük megszekvenálása után, úgy tűnik, hogy végre sikerül megtalálni a valódi helyüket: ez pedig - láss csodát - a csalánozók között van. Az eredményeket ismertető cikk oxfordi szerzői szerint ez ugyanakkor nem azért van, mert a csalánozók ősi formáját tükröznék (sőt, féregalkatuk ellenére, ezek nem kétoldalian szimmetrikus állatok, nincs idegrendszerük, érzékszerveik, vagy felismerhető bélcsatornájuk), inkább arról van szó, hogy egy virágállatok testfelépítése módosult a hosszú év milliók során, hogy egy fonálféregszerű (de nem fonálféreg) lény jöjjön létre.
Putnam, NH, Srivastava, M, Hellsten, U, Dirks, B, Chapman, J et al. (2007) Sea Anemone Genome Reveals Ancestral Eumetazoan Gene Repertoire and Genomic Organization. Science 317: 86-94.
Jiménez-Guri, E, Philippe, H, Okamura, B, Holland, PWH (2007) Buddenbrockia Is a Cnidarian Worm. Science 317: 116-118.