A közelmúltban az egyik magyarországi kreacionista műhely honlapjára felkerült egy iromány az ausztráliai homoki lábastyúk (Leipoa ocellata) költési szokásairól. A szöveg, amennyire megítélhető, a mozgalom frontembere, Tasi István által pár éve megjelentetett kreacionista pamflet, “A Természet IQ-ja” egyik fejezetének összefoglalását tartalmazza. A lényeg (amit emailben is eljuttattak hozzánk kreacionista “barátaink”) röviden: a homoki lábastyúk, a legtöbb madárfajjal ellentétben nem testmelegével kelti ki a tojásait, hanem költődombokat épít. Ezekben lebomló növényi anyagokat halmoz fel, és az állat a költődomb hőmérsékletét aktívan szabályozza, “hihetetlen pontossággal 34°C-on tartja; maximum egy fok eltérést engedélyez ehhez képest”. A nagyméretű tojásokból hosszabb keltési periódus után teljesen kifejlett fiókák kelnek ki, akik “legurulnak a költődombról, és rögtön bevetik magukat a környező bozótosba.” Szüleikkel nem találkoznak, mégis tudják majd, ha eljön az ideje, hogyan kell dombot építeni. Mivel nem képzelhető el, hogy a klasszikus tojáson kotló madár viselkedésből ez a hihetetlenül komplex és eredményes viselkedésforma lépésenként létrejöhetne, ez egy egyszerűsíthetetlenül összetett rendszer. Tehát Darwin tévedett, az evolúció hülyeség. Q.E.D.
A közelmúltban az egyik magyarországi kreacionista műhely honlapjára felkerült egy iromány az ausztráliai homoki lábastyúk (Leipoa ocellata) költési szokásairól. A szöveg, amennyire megítélhető, a mozgalom frontembere, Tasi István által pár éve megjelentetett kreacionista pamflet, “A Természet IQ-ja” egyik fejezetének összefoglalását tartalmazza. A lényeg (amit emailben is eljuttattak hozzánk kreacionista “barátaink”) röviden: a homoki lábastyúk, a legtöbb madárfajjal ellentétben nem testmelegével kelti ki a tojásait, hanem költődombokat épít. Ezekben lebomló növényi anyagokat halmoz fel, és az állat a költődomb hőmérsékletét aktívan szabályozza, “hihetetlen pontossággal 34°C-on tartja; maximum egy fok eltérést engedélyez ehhez képest”. A nagyméretű tojásokból hosszabb keltési periódus után teljesen kifejlett fiókák kelnek ki, akik “legurulnak a költődombról, és rögtön bevetik magukat a környező bozótosba.” Szüleikkel nem találkoznak, mégis tudják majd, ha eljön az ideje, hogyan kell dombot építeni. Mivel nem képzelhető el, hogy a klasszikus tojáson kotló madár viselkedésből ez a hihetetlenül komplex és eredményes viselkedésforma lépésenként létrejöhetne, ez egy egyszerűsíthetetlenül összetett rendszer. Tehát Darwin tévedett, az evolúció hülyeség. Q.E.D.
 A posztba beillesztve még megtalálhatjuk Tasi István pávák társaságában megejtett Attenborough imperszonációját is, és ha másért nem, ezért mindenképpen érdemes megtekinteni a művet. (Bár, elkötelezett Attenborough rajongókként mi* inkább az eredetit ajánlanánk - ha valaki siet, az tekerjen a 00:32:11-00:34:20 közti részhez).
A posztba beillesztve még megtalálhatjuk Tasi István pávák társaságában megejtett Attenborough imperszonációját is, és ha másért nem, ezért mindenképpen érdemes megtekinteni a művet. (Bár, elkötelezett Attenborough rajongókként mi* inkább az eredetit ajánlanánk - ha valaki siet, az tekerjen a 00:32:11-00:34:20 közti részhez).
A történettel (szokás szerint) csak néhány kisebb bökkenő akad. A viselkedésforma közel sem annyira hatékony, ahogy azt Tasi bemutatja (még a tutifixnek látszó 34+/-1°C sem igaz ebben a formában), az “egyszerűsíthetetlen” viselkedésforma számos eleme előfordul más madaraknál, és ami talán a legfontosabb, nem túl nehéz találunk valószínűsíthető és logikus evolúciós uta(ka)t a viselkedés kialakulásához. Vegyük akkor mindezeket sorba.
A közelmúltban az egyik magyarországi kreacionista műhely honlapjára felkerült egy iromány az ausztráliai homoki lábastyúk (Leipoa ocellata) költési szokásairól. A szöveg, amennyire megítélhető, a mozgalom frontembere, Tasi István által pár éve megjelentetett kreacionista pamflet, “A Természet IQ-ja” egyik fejezetének összefoglalását tartalmazza. A lényeg (amit emailben is eljuttattak hozzánk kreacionista “barátaink”) röviden: a homoki lábastyúk, a legtöbb madárfajjal ellentétben nem testmelegével kelti ki a tojásait, hanem költődombokat épít. Ezekben lebomló növényi anyagokat halmoz fel, és az állat a költődomb hőmérsékletét aktívan szabályozza, “hihetetlen pontossággal 34°C-on tartja; maximum egy fok eltérést engedélyez ehhez képest”. A nagyméretű tojásokból hosszabb keltési periódus után teljesen kifejlett fiókák kelnek ki, akik “legurulnak a költődombról, és rögtön bevetik magukat a környező bozótosba.” Szüleikkel nem találkoznak, mégis tudják majd, ha eljön az ideje, hogyan kell dombot építeni. Mivel nem képzelhető el, hogy a klasszikus tojáson kotló madár viselkedésből ez a hihetetlenül komplex és eredményes viselkedésforma lépésenként létrejöhetne, ez egy egyszerűsíthetetlenül összetett rendszer. Tehát Darwin tévedett, az evolúció hülyeség. Q.E.D.
A posztba beillesztve még megtalálhatjuk Tasi István pávák társaságában megejtett Attenborough imperszonációját is, és ha másért nem, ezért mindenképpen érdemes megtekinteni a művet. (Bár, elkötelezett Attenborough rajongókként mi* inkább az eredetit ajánlanánk - ha valaki siet, az tekerjen a 00:32:11-00:34:20 közti részhez).
A történettel (szokás szerint) csak néhány kisebb bökkenő akad. A viselkedésforma közel sem annyira hatékony, ahogy azt Tasi bemutatja (még a tutifixnek látszó 34+/-1°C sem igaz ebben a formában), az “egyszerűsíthetetlen” viselkedésforma számos eleme előfordul más madaraknál, és ami talán a legfontosabb, nem túl nehéz találunk valószínűsíthető és logikus evolúciós uta(ka)t a viselkedés kialakulásához. Vegyük akkor mindezeket sorba.
Tökéletes tervezés?
A homoki lábastyúk, már csak elterjedésénél fogva sem örvendezhet akkora kutatói figyelemnek, mint számos más madárfaj, ami meg is látszik a rendelkezésre álló irodalom viszonylagos szegénységén. Ugyanakkor az, ami fellelhető, kiegészítve az ausztrál kormány készítette (és irodalmi referenciákkal bőven ellátott) rövidke ismertetővel, már bőven elégnek tűnik a hirtelen támadt tudásszomj kielégítésére.
A madár életciklusáról szóló adatok különösen érdekesek, hiszen egy valóban tervezett rendszertől elvárnánk, hogy az nagy hatékonysággal működjön, vagyis a lábastyúk fiókák mortalitási aránya alacsony legyen. De nem az. A gondok már tojásrakáskor kezdődnek, ui. a tojások kb. 14,1%-a eleve megteremékenyítetlen. Ráadásul a megtermékenyítés mesze nem garantálja a kikelést: ezek a tökéletes „ösztönprogrammal” rendelkező madarak a fészek rendezgetése közben ugyanis rendszeresen megsértik az amúgy igen vékony héjjal rendelkező tojásokat. Olyannyira, hogy ez az ügyetlenség az embriókori mortalitás 4%-áért felelős, így Tasi István állítása, miszerint „a homoki lábastyúk az apai gondoskodás mintaképe”, egy legalábbis vitatható szubjektivitású állítás. Végül a költődomb tojásainak csak az 50-85%-a kel ki (sőt, ez az arány bizonyos időszakokban 50% alá eshet). Ám mindez semmi ahhoz képest, ami ezután jön: a csibék 80%-a az első hetekben elpusztul a ragadozók karmai közt, vagy metabolikus stressz miatt. Később a mortalitás csökken, de még mindig jelentős marad. Ezek a tényezők is hozzájárulhatnak ahhoz, hogy a faj jelenleg veszélyeztetett státuszú, így jövője és fennmaradása bizonytalan.
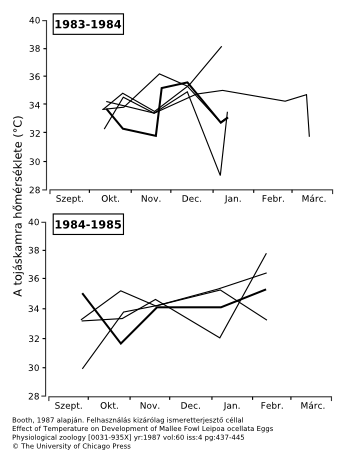 Mint említettük, a “34+/-1°C” is pontosításra szorul. A lábastyúk-költődombok belsejének átlagos hőmérséklete valóban 34°C, de azért egy foknál lényegesen nagyobb ingadozás mérhető. Konkrétan akár négy fokkal több vagy kevesebb is mérhető természetes körülmények között, akár több napon át is, a költési periódus során[1]. A “termosztát” tökéletes működését cáfolják azok a megfigyelések is, melyek szerint az esős periódusokban a hirtelen lezúduló csapadék nyomán akár a költődombban levő összes tojás elpusztulhat[2].
Mint említettük, a “34+/-1°C” is pontosításra szorul. A lábastyúk-költődombok belsejének átlagos hőmérséklete valóban 34°C, de azért egy foknál lényegesen nagyobb ingadozás mérhető. Konkrétan akár négy fokkal több vagy kevesebb is mérhető természetes körülmények között, akár több napon át is, a költési periódus során[1]. A “termosztát” tökéletes működését cáfolják azok a megfigyelések is, melyek szerint az esős periódusokban a hirtelen lezúduló csapadék nyomán akár a költődombban levő összes tojás elpusztulhat[2].
Ezen viselkedés további érdekessége, hogy az erre kapható madárfajok (nemcsak a Leipoa, de erről bővebben majd később) meglepően önfejűek: ha egyszer elkezdenek egy dombot építeni, akkor nagyon nehezen beszélhetőek le annak befejezéséről, ami a jelek szerint nem kevés bosszúságot okozhat az ausztrál hobbikertészeknek. (Ugyanakkor egy jó helyre épített komposzt-gödörrel a Leipoával rokon, szintén költődombot építő fajok hímjei lekenyerezhetők, és azt fogják továbbépíteni költődombbá.)
Egyszerűsíthetetlen komplexitás?
A viselkedés tehát komplex, de közel sem tökéletes. Na de vajon tényleg leegyszerűsíthetetlen-e? Azaz egyes elemeit fel lehet-e fedezni más fajok esetében? (Ugye ezek elvileg magukban nem működhetnek, csak az egész pakk egyben.)
Tasi maga felsorol néhány vizsgálandó elemet, fussunk ezeken át:
1. „A tojó részéről: Visszatérés rendszeres időközönként és a tojások lerakása a megfelelő helyre”.
A fekete rigótól a bíbicen át a kolibriig minden több tojást rakó madárfaj tojója pontosan ezt csinálja, ami a leírásban szerepel: rendszeres időközönként visszatér és a tojásait lerakja a megfelelő helyre, anélkül, hogy költődombot építene. Mivel ez lehetetlen, hiszen egy leegyszerűsíthetetlenül összetett viselkedés elemei önmagukban nem lehetnek működőképesek, sem a rigó, sem a bíbic, sem a kolibri nem létezhet.
2.„A hím részéről: A domb alapanyagairól és struktúrájáról szóló tudás”.
Ugyan miért kellene a lábastyúknak bármit is tudnia a domb alapanyagairól? Az értelmes tervezés szónoka ellentmondásban van saját magával, hisz másutt azt írja, hogy a költődomb építése „veleszületett ösztönprogram” eredménye, és hogy ezt a tevékenység nem írható a madarak kognitív képességeinek a számlájára, mert az messze meghaladná a madarak „gondolkodási kapacitását”. Most ugyanakkor azt várja el a madártól, hogy az alapanyagokról való tudással rendelkezzen. Ezek szerint a hálószövő pókoknak, amelyek szintén „ösztönprogram” szerint építik a hálójukat, tisztában kellene lenniük a trigonometria szabályaival és a selymet felépítő komplex proteinstruktúrákkal? Vagy mi csak úgy használhatjuk sikeresen a mobiltelefonunkat, ha tisztában vagyunk az integrált áramkörök működési elvével? Teljesen nyilvánvaló, hogy ez a pont értelmetlen: a madár a fészekdomb struktúrájáról való tudás nélkül is használhatja azt, akárcsak a pók a hálóját, vagy mi a telefonunkat. Ráadásul az sem mellékes a témában, hogy rokon és szintén költődombépítő fajok gyakran emberi hulladékot (italos dobozokat, foci labdát, kanalakat) is beleszerkesztenek az építményeikbe, ami szintén jól demonstrálja, hogy ez a “tudás” egy igencsak homályos és plasztikus ösztönhalmaz.
3. “Hőérzékelő szerv”
Nem világos például, hogy Tasi István miért gondolja, hogy a hőérzékelés a lábastyúk egyedülálló sajátossága, ami csak a dombépítéssel kapcsolatban jelent meg. Mi az alapja annak a feltételezésnek, hogy más madarak nem rendelkezhettek hőérzékelésre alkalmas receptorokkal? A valóság az, hogy termoreceptorok jelenlétét a csőr illetve a nyelv területéről már több alkalommal is leírták más fajokban[3].
4. “Az állandó hőmérséklet biztosításához szükséges ösztönös tevékenységek.”
Más fajok is nagy gondot fordítanak pl. a tojások ki-be takargatására, költődomb nélkül is. Földön fészkelő széki lilék (Charadrius alexandriunus) például napszakonként változtatják a tojásokat takaró fészekanyag mennyiségét[4] (úgy tűnik ők is kiváló időjósok), és ha a kutatók kísérletesen megnövelték vagy lecsökkentették fészkeikben a fészekanyag mennyiségét, a szülők hamar visszaállították[5] az eredeti állapotot, akárcsak tették azt Attenborough filmjében is a lábastyúkok. A forró területen fészkelő széki lilék ráadásul a külső hőmérséklet függvényében változtatják kotlási viselkedésüket[6]. Napközben, a rekkenő hőségben (a hőmérséklet meghaladhatja az 50˚C-t) a pár valamelyik tagja mindig a fészken ül, hisz a magára hagyott tojások hamar elpusztulnának a hőségben, míg a hűvösebb reggeli és esti órákban hosszabb időre mindketten elhagyják a fészket.
Talán még érdekesebb a krokodilmadár (Pluvianus aegyptius) esete. Ez a madár a tojásait homokkal borítja be, és ezen ülve kotlik, majd éjszaka kitakarja azokat[7]; tehát a tojások homokkal való beborítása sem egyedülálló a lábastyúknál.
5. “A csibe részéről: Ösztönösen helyes viselkedés, hogy mi a teendő a tojásból való kibújás után.”
6. “Születéstől adott ösztönök a későbbi párválasztáshoz és utódgondozáshoz.”
7. “Megfelelő anatómiai fejlettség, hogy legyen erejük kiásni magukat a dombból, és hogy egyedül életben maradhassanak.”
Korábban már említettük, hogy a lábastyúk embriók és fiókák mortalitása igen magas. Itt a fiókakori mortalitás egy eddig nem említett elemére hívnánk fel a figyelmet. A kikelt fiókák 2.9%-a ugyanis sosem jut a felszínre, hanem elpusztul a költődombban. Nehéz tudni, hogy ez azért történik, mert a fiókák viselkedése valamiért nem megfelelő (5. pont) vagy mert pl. a fiókák nem elég fejlettek vagy erősek ahhoz, hogy kiássák magukat a dombból (7. pont). Egy másik Megapodiidae fajon végzett vizsgálat szerint[12] a költődomb hőmérsékleti ingadozása hatással van a fiókák keléskori súlyára, viszont nincs hatással a keléskori testméretre. Bár nincs tudomásunk hasonló vizsgálatról a Leipoa-nál, okkal feltételezhetjük, hogy a költődomb hőmérsékletének ingadozása náluk is hasonló hatással lehet a fiókák fejlődésére. Az azonos méretű, de kisebb súllyal rendelkező (alacsonyabb hőmérsékleten kelt) fiókák tehát kevesebb zsírtartalékkal rendelkeznek, rosszabb fizikai kondícióban vannak, így valószínűsíthető ez az oka annak, hogy egy részük nem jut a felszínre, hanem megfullad a fészekben. A rendszer tehát itt sem tökéletes, hanem a fiókák kelési és felszínre jutási sikere, akárcsak a lábastyúk viselkedésének bármely eleme változatos, és a természetes szelekció azokat a változatokat részesíti előnyben, amelyek az egyedeknek a legnagyobb túlélést és szaporodási sikert biztosítják.
Bár hamarosan belevágunk a filogenetikai lecsóba, de az már előre kívánkozik, amolyan teaserként, hogy a lábastyúk a Tyúkformájúak (Galliformes) közé tartozik. Ennek a rendnek az egyik fontos jellegzetessége a csibék nagyfokú önállósága, mert átlagosan órákkal a születés után képesek a szüleikkel együtt mozogni és táplálékot keresni. (Egyébként az ősi madárcsoportokra ez általában jellemző, a magatehetetlenül és meztelenül tátogó madárfiókákat a modernebb csoportok esetében találjuk.) Ugyan a Tyúkformájúak között a lábastyúk kétségtelenül az önállóság extrém példája, de nem lóg kia a mezőnyből, hanem egy folyamatos spektrum végén helyezkedik el[8]. Például egy kiskacsa nem sokkal magatehetelenebb nála, hiszen születés után azonnal mozogni, úszni és táplálkozni tud. (A többi referencia mellett bátran ajánljuk itt Szutyejev meséit is ;-))).
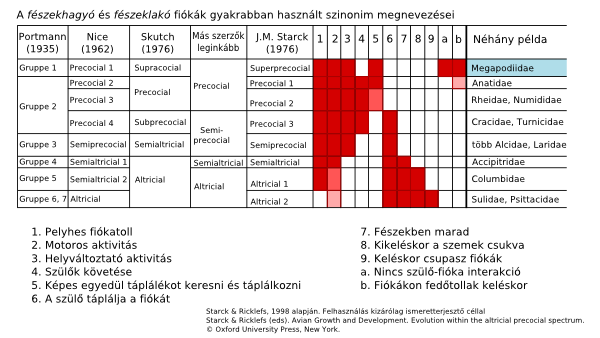
Láthatjuk, hogy a komplex viselkedésforma egyes elemei felfedezhetőek más fajokban is, és őszintén szólva egyáltalán nem ütközik semmilyen különösebb logikai nehézségbe egy olyan forgatókönyv felállítása, ami szerint a lábastyúk mai – húzzuk alá – tökéletlen viselkedése lépésről lépésre alakuljon ki.
De mielőtt a viselkedés kialakulására egy lehetséges útvonalat felvázolnánk, lássuk csak, mit tudunk a szóban forgó faj rendszertani besorolásáról és evolúciójáról.
A Megapodiidae család rendszertana és evolúciója
A homoki lábastyúk a Megapodiidae családba tartozik számos rokonával, akik közül sokan hasonló költődombokat építenek, de mindegyik kivétel nélkül “tojásülő” program nélkül kelti ki fiókáját. Tasi István szövege picit homályos a téren, hogy ő most nincs tisztában ezeknek a rokon fajoknak a létével, vagy pedig a Leipoa az egész csoport költődombos fajainak mintapéldánya akar lenni - az egyszerűség kedvéért most az utóbbit feltételezzük, bár nem vagyunk biztosak, hogy optimizmusunknak van alapja. Az idetartozó fajok egyaránt erőteljes kapirgáló lábakkal, s aránylag rövid, lekerekített szárnyakkal rendelkeznek. A legkisebb termetű fajaik képesek aránylag gyors és nagyobb távolságra történő repülésre (a történelmi időkben még nagytermetű és már röpképtelen fajaik is éltek).
Legkülönlegesebb és a madarak körében egyedülálló tulajdonságuk, hogy a tojásaikat nem a szülők költik ki, s általában nem is más fajjal költetik ki, mint például a kakukk (bár néhány parazita faj azért köztük is akad[9]). A testméretükhöz képest aránylag nagy tojásukat (amely a nőstény testének 15-20%-át is elérheti!) napsütötte homok dűnékbe ásott üregbe, gejzírek közelében, a nap által felmelegített talaj üregeibe, vagy éppen a több tojó által összekapart levélrakásba, illetve a Leipoa esetében homok és vegetáció kombinációjából készült költődombba helyezik el. A levélrakás és költődomb esetében az állatok időnként ellenőrzik is a domb hőfokát és ennek megfelelően rendezik át a fészekaljat. A többi költési stratégiát követő fajok viszont rendszerint teljesen magára hagyják a lerakott tojást, amit a környezet hője kelti ki (hasonlóan egyes hüllők viselkedéséhez).
A fióka tojásban való fejlődése igen hosszú ideig tart, ami eléri sőt meghaladja a 90 napot is, amihez a nagyméretű tojás szikanyagtartalma biztosítj az ellátást. Viszont a kikelt fióka már teljesen ki van tollasodva, saját maga ássa ki magát a költőhelyéből és rögtön képes önálló életet kezdeni.
A Megapodiidae családba jelenleg 7 nemzetség 22 faja tartozik. Kivétel nélkül Ausztráliában és a környékbeli kontinentális eredetű szigeteken élnek (Új Guinea, Új Kaledónia, Új Zéland, stb). Számos fajuk már a korábban kihalt, feltehetőleg több esetben vadászat, illetve betelepített ragadozók miatt halt.
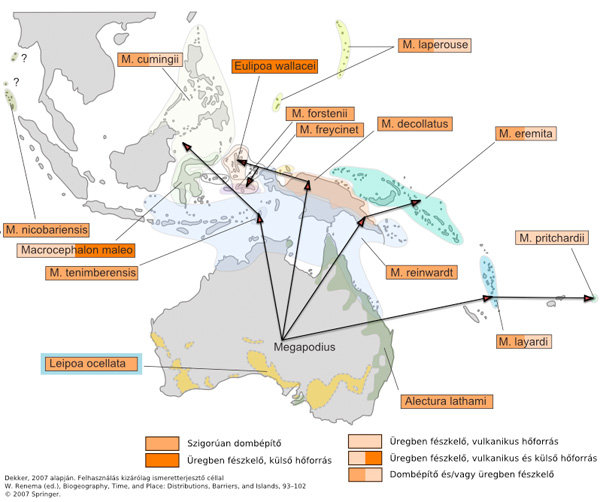
A család legkorábbi képviselői már az Eocénban megjelennek (35-45 millió évvel ezelőtt (MÉ)) és a korai miocénnel bezárólag (15-20 MÉ) nemcsak a mai élőhelyükről, hanem Dél-Amerikából és Európából is ismertek fosszilis fajokkal. Ezeket egy új családba, az egyik legjelentősebb franciaországi lelőhely (Quercy) után elnevezett Quercymegapodiidae csoportba sorolták, s ebbe két nemzetség (Quercymegapodius és Ameripodius) tartozik 2-2 fosszilis fajjal. Pár éve Romániából, a Kárpát-medence külső pereméről, szintén eocén rétegből került elő egy új faj szinte teljes csontváz lenyomata, ami jelenleg leírás alatt áll.
A madarak klasszikus és genetikai rendszerezése egyaránt a legősibb szárazföldi valódi madarak (Neornithes) közé sorolja a Tyúkalkatúakat (Galliformes), illetve a genetikai taxonómiában a Gallo-Anseres csoportot.
Hol alakulhatott ki (földrajzilag) a család? Az a tény, hogy a család képviselői már a harmadkor elejétől fellelhetők és ráadásul bizonyítottan két, a mai élőhelytől eltérő kontinensen is, ez két lehetőség taglalására nyit utat:
Vagy még a Pangea feldarabolódása előtt már kialakult az ősük és a feldarabolódó őskontinens egymástól eltávolodó részein önállóan fejlődtek tovább a család fajai, vagy csak a mezozoikum végén jelentek meg Laurasiában, majd egyik csoportjuk É-Amerikán keresztül jutott el Dél Amerikába, míg a másik pedig északnyugat felől került Ausztráliába és a környező nagyobb szigetekre.
Ősföldrajzi és madárevolúciós meggondolások alapján az előbbi változatot tartjuk valószínűbbnek.
Variációk költődombépítés-evolúcióra
Ennyi felvezetés után lássuk a medvét, vagyis milyen evolúciós utakat feltételezhetünk a költődomb-építő viselkedés kialakulására. (Rögtön kettővel is szolgálunk, amelyek minimális irodalmazás után, eléggé kézenfekvőnek tűnnek.)
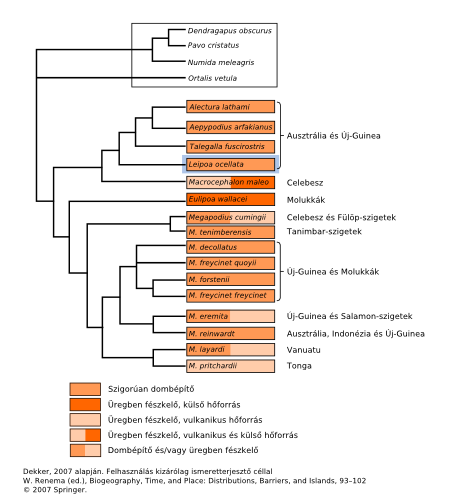 Ha a család filogenetikai fáján feltüntetjük a költési szokásokat, akkor azt figyelhetjük meg, hogy két nagy csoport különül el[10]. Mind a kettőben vannak költődomb építők (mound builder), ugyanakkor van ahol ezt a viselkedésformát egyszerű üreg készítés (burrow nesting) előfordulása is kíséri, és van, ahol kizárólag az utóbbi figyelhető meg.
Ha a család filogenetikai fáján feltüntetjük a költési szokásokat, akkor azt figyelhetjük meg, hogy két nagy csoport különül el[10]. Mind a kettőben vannak költődomb építők (mound builder), ugyanakkor van ahol ezt a viselkedésformát egyszerű üreg készítés (burrow nesting) előfordulása is kíséri, és van, ahol kizárólag az utóbbi figyelhető meg.
Mivel mindkét csoport legősibb formái (Macrocephalon maleo és Eulipoa wallacei) esetében nincs költődomb építés, az egyik elmélet az, hogy az üregásás volt a csoport ősibb viselkedésformája, és ebből aztán párhuzamosan, több helyen is kialakult a dombkészítés. Annak egyszerűbb formáiban, melegebb éghajlaton, ez mindössze szerves-anyag felhalmozást jelentett (vagyis egyszerűen az aljzaton levő rothadó levelekbe való tojás helyett az állatok maguk készítettek egy kupacot), míg a szárazabb területeken élő Leipoa ezt fokozta tovább, kidolgozva egy homokkal egybekötött kombinációt. (Megjegyzendő, hogy a Leipoa az egyetlen száraz területen élő faj.)
A másik, párhuzamos eszmefuttatás abból indul ki, hogy a pusztán természetes sugárzási forrásokra hagyott tojáskeltetés elsősorban melegebb égövön megvalósítható csak, így, ha elfogadjuk, hogy a család eredete mérsékelt égövön volt, akkor az aktív(abb) hőtermeltetés tűnik az ősibb tulajdonságnak, és az erről való “leszokást” csak a meleg területeken élő fajok engedhették meg maguknak. Azaz a költődombon alapuló költési forma, ergo a költődombot létrehozó viselkedés alakult ki előbb.
Hogy melyik elképzelés helytálló, azt nehéz eldönteni, ugyanis a történelmi korokban létezett Megapoda ősök költési szokásairól nagyon keveset tudunk, és kérdés, hogy ez a helyzet valaha meg fog-e változni.
Persze mindezzel még mindig nem válaszoltuk meg Tasi legnagyobb problémáját, hogy miként alakulhatott ki ez a “nem kotlós” magatartásforma a többi madárra jellemző tojáson ülősből.
Ha az első elméletet próbáljuk végigkövetni (vagyis az “elássuk a tojásokat, majd odébbállunk” volt előbb), akkor leginkább az érdemel említést, hogy ez a viselkedés igen gyakori hüllőknél. Akár viselkedési atavizmusról is beszélhetünk: az ősben élő viselkedési ösztönök újra bekapcsolnak, és annak megfelelően tevékenykedik majd az állat. Az sem zárható ki persze, hogy nem atavizmusról, hanem adaptációról van szó: nyilvánvaló, hogy előnyös az a lehetőség, hogy szülők helyett a környezeti közeg melegére lehessen bízni a tojások kiköltését. Ezáltal csökkenhet a ragadozók általi fenyegetettség, valamint megszűnt az inkubációs kapacitás okozta korlát a fészekaljméretben (magyarán egy párnak több tojása lehet, mint amennyit maguktól ki tudnának kelteni)
A második elmélet részletezéséhez abból indulunk ki, (amit előzőleg már tisztáztunk), hogy a lábastyúkok viselkedése messze nem egyszerűsíthetetlen, épp ellenkezőleg, viselkedésük számos eleme megtalálható más fajokban is, amelyek nem építenek költődombot. Így könnyen elképzelhető, hogy ezek a Megapodiidae leszármazási ághoz vezető fajokban is megtalálhatóak voltak. A tinamufélék között például gyakori jelenség, hogy amikor elhagyják a fészket, azokat levelekkel takarják be[11]. Amennyiben ezek a levelek akár csak kismértékben is megakadályozták a tojások kihűlését, a fészek tulajdonosának több ideje juthatott táplálkozásra, párosodásra, stb. ezáltal több utódja lehetett, mint az ezt a viselkedést nem tanúsító egyedeknek (hiszen a kotlásra fordított idő a többi tevékenység rovására megy). Ennek következtében a viselkedés elterjedhetett a populációban. Meg kell jegyezni, hogy a tinamuk elsősorban a tojások elrejtése miatt takarják be a tojásaikat, és lehetséges, hogy a Megapoda-k evolúciójának kezdetén a tojások betakarása is ezt a célt szolgálta, és nem a tojások szigetelését, és ez utóbbi funkció csak másodlagosan kapott szerepet. Elképzelhető, hogy hamar megjelent egy újabb viselkedésforma, ami nem csak levelekkel takarta be a tojásokat, de arra némi földet is kapart (közismert, hogy a Megapodák rokonságába (velük egy rendbe) tartozó tyúkfélékre jellemző a kapirgálás, így a viselkedés megjelenése a fészek körül sem valószínűtlen). A korábban említett krokodilmadár szintén homokkal borítja be a tojásait. Ha tehát a levelekkel betakart tojások homokkal való elfedése tovább növelte a tojások termikus stabilitását vagy azok védettségét a ragadozók elől, akkor ez előnyhöz juttathatta az ezt a viselkedést mutató egyedeket.
Összességében, valóban számos érdekes kérdés maradt még nyitva a lábastyúkokkal kapcsolatban, de mindez nem teszi lehetetlenné, hogy felvázoljunk egy, a “a természettudományos ismeretekkel összhangban álló magyarázatot” egy valójában koránt sem tudományos igénnyel felvetett problémára. (Mivel Tasi István nem jelöli meg az általa áttekintett irodalmi forrásokat, kíváncsiak lennénk, pontosan milyen műveket használt fel a könyve elkészítésekor, és az általunk érintett irodalmat miért nem lelte meg kutakodása közben, vagy, ha meglelte, miért nem foglalta bele az írásába...?)
Tovább, tovább
Végül, mivel a lábastyúk gondolatköre láthatóan megfogta a Védikus Tudományok Kutatóközpontjában dolgozók fantáziáját, ajánlanánk néhány kutatási vonalat, amit követve érdekes eredményekre bukkanhatnának:
- Megfigyelések szerint óriási változatosság van az egyes párok viselkedése között abban, hogy a hím és a tojó mennyit dolgozik a költődombon. (Tasi István állításával ellentétben ui. a tojó gyakran gondozza a költődombot.) Ugyanez a változatosság jellemzi a kelési sikert, a tojások számát, stb. Nem világos számunkra, hogy az atyai tervezettség szerint mi állhat e változatosság hátterében. Ha a madarak viselkedése egy felsőbbrendű intelligencia precíz mérnöki munkájának eredménye, mi okozza az egyedek közötti eltéréseket? Ráadásul maguk a tervezettség hívei állítják, hogy a viselkedés teljesen veleszületett, „program”-szerűen kódolt (amivel ugyan lehet vitatkozni, de most ettől tekintsünk el). Miért nem követi minden egyed ugyanazt a tervet? Vagy ezek szerint több program is létezik?
- Melyik párnak lesz több utódja, amelyik 3 vagy amelyik 33 tojást gondoz a költődombjában (ez a megfigyelt változatosság a fészekalj méretben)? Mi lesz a a populáció összetételére nézve a következménye annak, ha a fészekaljméretben tapasztalható különbség több generáción keresztül megmarad?
- Mi az oka annak, hogy a ma élő Megapodiidae fajok szaporodási rendszere igen változatos? Mivel magyarázható, hogy egyes fajok poligínek, mások monogámok?
- Mi az oka annak, hogy a Megapoda fajok kizárólag az ausztráliai és óceániai régióban fordulnak elő?
- Mi az oka annak, hogy egyes Megapoda fajok (pl. Aepypodius bruijnii) populációmérete mindössze néhány száz egyedre zsugorodott, míg egyes fajok pedig kifejezetten abundánsak (pl. Alectura lathami)?
- Mi az oka annak, hogy a Progura gallinacea nagyméretű Megapoda faj kipusztult? Értelmezhető-e ez úgy, hogy az ő „ösztönprogramja” nem volt tökéletes?
- Végül, egyetértve Tasi Istvánnal abban, hogy az evolúciós genetikai változások genetikai hátterének feltárása igen fontos, javasolnánk, hogy a Kutatóközpont fogjon bele a Leipoa genom megszekvenálásába. A szekvenálási költségek csökkenésével ez ma már nem elképzelhetetlen (ráadásul a csirke genom referenciaként nagyban megkönnyíti a műveletet) és szépen beleillene a tízezer faj genomjának feltárását megcélzó Genome10K programba. Kutatásukhoz sok sikert kívánunk!
*(A poszt a következő négy szerző közös munkáján alapul: Ambrus Géza Gergely, Kessler Jenő (primavis), Lendvai Ádám és Varga Máté (dolphin).)
[1] Booth DT (1987) Effect of Temperature on Development of Mallee Fowl Leipoa ocellata Eggs. Physiological Zoology 60: 437-445.
[2] Brickhill (1987) Breeding success of Malleefowl Leipoa ocellata in Central New South Wales. Emu 87: 42-45.
[3] Whittow (ed) 2000 Sturkie’s Avian Physiologym 5th edition.
[4] Szentirmai & Székely (2004) Diurnal variation in nest material use by the Kentish Plover Charadrius alexandrinus. Ibis 146, 535–537
[5] Szentirmai & Székely (2002) Do Kentish plovers regulate the amount of their nest material? An experimental test. Behaviour, 139: 847-859.
[6] AlRashidi et al. (2010) The influence of a hot environment on parental cooperation of a ground-nesting shorebird, the Kentish plover Charadrius alexandrinus. Frontiers in Zoology, 7: 1.
[7] Seymour & Ackerman (1980) Adaptations to underground nesting in birds and reptiles. American Zoologist, 20: 437-447.
[8] Avian Growth and Development. Evolution within the altricial precocial spectrum. J. M. Starck and R. E. Ricklefs (eds). Oxford University Press, New York, 1998
[9] Jones & Birks (1992) Megapodes: Recent ideas on origins, adaptations and reproduction. Trends in Ecology and Evolution, 7: 88-91
[10] Dekker: Distribution and Speciation of Megapodes (Megapodiidae) and Subsequent Development of their Breeding Behaviour (in Biogeography, time and place:distributions, barriers and islands - Renema W (ed) - Springer, 2007)
[11] Hoyo et al. Handbook of the Birds of the World. Volume 1.
[12] Seymour 1995 Thermochimia Acta 250: 319-328