A közelmúltban közzétett Human Genome Diversity Project (HGDP) első eredményei világrengetőnek nem igazán nevezehetőek: lényegében minden fontosabb eredmény már ilyen-olyan formában ismert volt, vagy mitokondriális, vagy X és Y kromoszómás vizsgálatokból, vagy kisebb elemű egyéb genetikai felmérésekből.
A közelmúltban közzétett Human Genome Diversity Project (HGDP) első eredményei világrengetőnek nem igazán nevezehetőek: lényegében minden fontosabb eredmény már ilyen-olyan formában ismert volt, vagy mitokondriális, vagy X és Y kromoszómás vizsgálatokból, vagy kisebb elemű egyéb genetikai felmérésekből.
Mégis, az 51 populáció, 938 (rokonsági viszonyban nem álló) egyedén, 650,000 egyszerű nukleotida polimorfizmus (SNP) figyelembevételével végzett vizsgálat már a puszta adatmennyiség miatt is jelentős. És ha korábbi ismereteink megerősítést nyernek, hát annál jobb ;-).
A közelmúltban közzétett Human Genome Diversity Project (HGDP) első eredményei világrengetőnek nem igazán nevezehetőek: lényegében minden fontosabb eredmény már ilyen-olyan formában ismert volt, vagy mitokondriális, vagy X és Y kromoszómás vizsgálatokból, vagy kisebb elemű egyéb genetikai felmérésekből.
Mégis, az 51 populáció, 938 (rokonsági viszonyban nem álló) egyedén, 650,000 egyszerű nukleotida polimorfizmus (SNP) figyelembevételével végzett vizsgálat már a puszta adatmennyiség miatt is jelentős. És ha korábbi ismereteink megerősítést nyernek, hát annál jobb ;-).
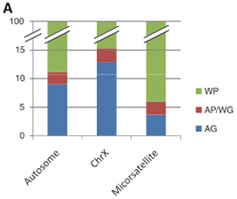 Az egyik ilyen ismerős eredmény az emberek közti genetikai sokszínűség eloszlását mutatja. A legtöbb különbség mindig egy-egy populáció egyes egyedei (within population - WP) közt figyelhető meg - a teljes sokféleség közel 80%-a fér ebbe a kategóriába. (Magyarán közted, nyájas olvasó, és átlagos szomszédod közt a genetikai különbség kb. a közted és egy eldugott indonéziai törsz random tagja közti különbség 80%-át teszi ki.) Az egyes földrajzi csoportok közötti eltérések (among geographical groups - AG) a sokféleség 4-12%-áról tehetnek, míg az egyes csoportok különböző populációi közti eltérés (among-population, within group - AP/WG) - pl. soproniak és szegediek - teszi ki a fennmaradó néhány százalékot.
Az egyik ilyen ismerős eredmény az emberek közti genetikai sokszínűség eloszlását mutatja. A legtöbb különbség mindig egy-egy populáció egyes egyedei (within population - WP) közt figyelhető meg - a teljes sokféleség közel 80%-a fér ebbe a kategóriába. (Magyarán közted, nyájas olvasó, és átlagos szomszédod közt a genetikai különbség kb. a közted és egy eldugott indonéziai törsz random tagja közti különbség 80%-át teszi ki.) Az egyes földrajzi csoportok közötti eltérések (among geographical groups - AG) a sokféleség 4-12%-áról tehetnek, míg az egyes csoportok különböző populációi közti eltérés (among-population, within group - AP/WG) - pl. soproniak és szegediek - teszi ki a fennmaradó néhány százalékot.
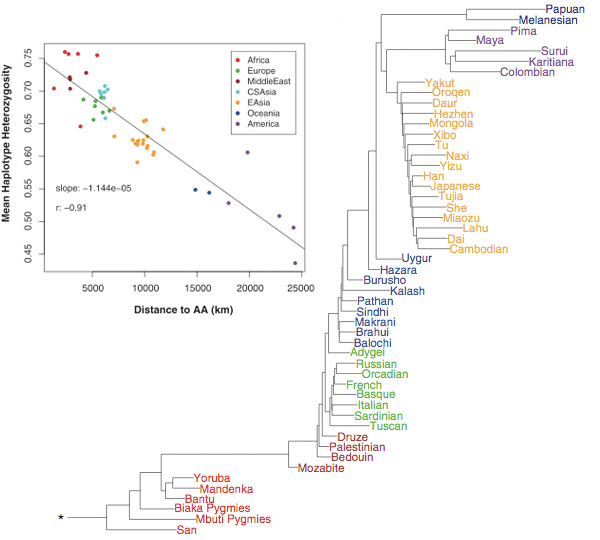
A másik (újra)felismerés az, hogy a heterozigozitás (vagyis egyes géneknek hány különböző formája, allélja fordítottan arányos az Afrikától való távolsággal (az alábbi ábrán a "Distance to AA" az Addis Abebától való távolságra vonatkozik). (Ez egyébként nemcsak genetikai de morfológiai variációra is igaz, ami szintén nem meglepő, hiszen a kettő nem független egymástól.) Ez a fajta géneloszlás pedig remekül magyarázható a Homo sapiens afrikai eredetével: a kezdeti közösség pár tagja távolabbra települt, de mivel nem hordozták magukban az anya populációban fellelhető összes allélvariációt, az új csoport genetikailag már csak kevésbé lesz genetikailag sokszínű. A folyamat ismétlődése pedig a Föld belakásán túl, épp a fent leírt genetikai mintázatot hozza majd létre.
A fentiekhez még hozzáadhatjuk a genetikai különbségek alapján szerkesztett filogenetikai fát is, amin egyrészt az egyes kontinenseken élő népcsoportok jól látható elkülönülése, másrészt az egyes nagyobb ágak közti reláció épp az Afrikából eredő, a Közel-Keleten Európa és Ázsia felé szétváló, majd Ázsiából Amerikába tartó vándorlási útvonalat valószínűsíti.
(De, bármennyire is vonzó a tétet az előbb emlegetett közelmúlti afrikai eredet elméletre tenni, az objektivitás azt kívánja, hogy azt is leszögezzük, bár a földrajzi távolság, heterozigozitás és genetikai távolság közti korreláció nagyon is valós, létezhetnek alternatív magyarázatok is. Ezeket boncolgatja John Hawks is a blogjában: nagyvonalakban arról van szó, hogy a késői pleisztocénben megfigyelt, afrikai eredetű génáramlás azt is simán jelentheti, hogy a pleisztocén során több hullámban vándoroltak ki az emberek Afrikából és az egyes vándorlások "genetikai nyoma" egymásra rakódott, s ezt érzékeljük ma. Laikus szem számára ez gyakorlatilag ugyanannak tűnhet, mint amit először is írtam, de van egy lényeges különbség: utóbbi esetben a Föld egyes területein élő emberi populációk úgy fogadhatták be az új géneket, hogy közben nem adták ét teljesen a helyüket az újonnan jötteknek. Nem kerülgetve tovább a kását: arról van szó, hogy az afrikai eredet alternatívájaként számontartott "multiregionális elmélet" (amely szerint a Föld különböző pontjain élő H. erectus populációk párhuzamosan alakultak át H. sapiens-é) nem teljesen halott, hanem elképzelhető, hogy a valóság a két elmélet hibridjében keresendő. Persze ez is egy teljes jogú elmélet, ami nem feltétlenül igaz, de a mai ismeretek szerint nem is feltétlenül hamis, pedig sokan úgy kezelik.)
Ha végül vigyázó szemeinket az európai adatokra tereljük, akkor két érdekességre figyelhetünk fel. Egyrészt, hogy a már "klasszikusnak" számító bőr-, szem- és hajszín, valamint immunrendszerhez kapcsolódó változásokon túl, nagyon sok új génvariáns a hideghez való alkalmazkodás jegyeit viseli magán. Elsősorban az anyagcseréhez kötődő génekről van szó, amelyek arra "idomultak" Európában, hogy hatékonyabban biztosítsák az állandó testhőmérsékletet, ami sok esetben a cukrok és zsírok potens feldolgozásában nyilvánul meg. Ez azonban azzal is járt bizonyos gének esetében, hogy a mára elterjedt variánsaik napjaink táplálékgazdag környezetében a cukorbetegségre való hajlamot növelik meg, azaz az egykori áldás mára átokká változott (igaz, más gének esetében az európai-allélok pont hogy csökkentik a diabetes hajlamot, szóval ki tudja).
A másik "európai" érdekesség, hogy úgy tűnik több "hibás" allél fedezhető fel bennünk, mint az afrikaiakban. A különbség kicsi - és sokak szerint nem is feltétlenül valós -, de ha igaz, akkor a legvalószínűbb magyarázata arra vezethető vissza, hogy Európa betelepítése viszonylag a közelmúltban következett be, egy relatíve kis méretű "telepes" közösség által. Amint a belakott terület nagyságával arányos a populáció mérete is növekedni kezdett, törvényszerűen új mutációk jelentek meg. Ezek többsége ilyen-olyan mértékben káros volt, de az eltelt idő rövidsége miatt nem tudta a természetes szelekció "kitakarítani" őket, a kontinens kollektív genetikai állományából.
(Már persze, ha valóban "káros" allélokról van szó. Nem szabad elfelejtkezni arról, hogy itt egy statisztikai megközelítésről van szó, ahol pusztán az tesz egy-egy genetikai variációt "károssá", hogy egy-egy gén olyan, nagyon konzervatív régiójába esik, ami sok faj között kvázi azonos. Ez valóban azt jelöli, hogy a régiót nehéz úgy változtatni, hogy az ne tegyen be a kódolt fehérje működésének, de azért funkcionális tesztek hiányában csak valószínűsíteni lehet egy ide beékelődő változás káros, biztosan tudni nem.)
Li JZ, Absher DM, Tang H, Southwick AM, Casto AM, et al. (2008) Worldwide human relationships inferred from genome-wide patterns of variation. Science 319: 1100-1104.
Jakobsson M, Scholz SW, Scheet P, Gibbs JR, VanLiere JM, et al. (2008) Genotype, haplotype and copy-number variation in worldwide human populations. Nature 451: 998-1003.
Lohmueller KE, Indap AR, Schmidt S, Boyko AR, Hernandez RD, et al. (2008) Proportionally more deleterious genetic variation in European than in African populations. Nature 451: 994-7.
Hancock AM, Witonsky DB, Gordon AS, Eshel G, Pritchard JK, et al. (2008) Adaptations to Climate in Candidate Genes for Common Metabolic Disorders. PLoS Genet 4(2): e32 doi:10.1371/journal.pgen.0040032