 Múlt héten a Magyar Távirati Iroda ügyeletes Lejter Jakabja igazán elemében lehetett, hiszen sikerült totálisan dezinformálnia, legalább egy hírecske erejéig, a teljes magyar online újságírást. "Az evolúció tüntette el az emberi péniszcsontot", hirdette a cím mindenfele, és hát nyilván dőlt a kattintásszám, a szerkesztő meg a marketing osztály elégedett volt, az pedig, hogy a hír ebben a formában légből kapott, kit érdekel...
Múlt héten a Magyar Távirati Iroda ügyeletes Lejter Jakabja igazán elemében lehetett, hiszen sikerült totálisan dezinformálnia, legalább egy hírecske erejéig, a teljes magyar online újságírást. "Az evolúció tüntette el az emberi péniszcsontot", hirdette a cím mindenfele, és hát nyilván dőlt a kattintásszám, a szerkesztő meg a marketing osztály elégedett volt, az pedig, hogy a hír ebben a formában légből kapott, kit érdekel...
Ugyanis az eredeti cikkben az emberekből valóban hiányzó péniszcsontról, a baculumról, szó egy szál sem esik. Helyette viszont van egy érdekes felvetés a pénisztüskék eltűnésének evolúciós okairól fajunkban - és csak azért felvetés, mert, mint végül látni fogjuk, a blogokban működő peer-review 2.0 néhány fontos és kényelmetlen kérdéssel állt elő a cikk kapcsán (amelyek, egyébként nem a kísérleti anyagot vitatják, hanem annak interpretációját, de ne rohanjunk ennyire előre).
A Nature-ben megjelent cikk fő témája olyan ember-specifikus deléciók (hCONDELs) vizsgálata, amelyeknek megfelelő DNS szakaszok például a nagyon közeli rokon csimpánzokban nemcsak, hogy jelen vannak, de igen nagyfokú konzervációt mutatnak más emlősök genomjával.
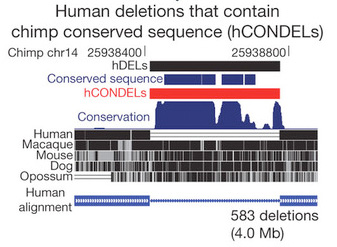 Ezeknek az ultrakonzervált szekvenciáknak a közös jellemzője, hogy nem fehérjekódoló régiókban vannak, mégis, gyakran jobban hasonlítanak egymásra, mint a közelükben levő fehérjekódoló szekvenciák, amelyek köztudottan lassabban változnak, mint a genom többi része. Mai feltételezések szerint ezeknek az elemeknek szabályozó szerepe van és ennek részben az az alapja, hogy ha ilyen szekvenciákat használva ún. riportergének meghajtására (amelyek, pl. zöld fluoreszcens fehérjét kódolnak), akkor a riporter-fehérje gyakran ugyanott fejeződik ki, mint ahol az ultrakonzervált szekvencia közelében levő gén. Vagyis, szól a cikkben is követett logika, ha ezek a szekvenciák hiányoznak, akkor annak valami szabályozási következménye kell legyen.
Ezeknek az ultrakonzervált szekvenciáknak a közös jellemzője, hogy nem fehérjekódoló régiókban vannak, mégis, gyakran jobban hasonlítanak egymásra, mint a közelükben levő fehérjekódoló szekvenciák, amelyek köztudottan lassabban változnak, mint a genom többi része. Mai feltételezések szerint ezeknek az elemeknek szabályozó szerepe van és ennek részben az az alapja, hogy ha ilyen szekvenciákat használva ún. riportergének meghajtására (amelyek, pl. zöld fluoreszcens fehérjét kódolnak), akkor a riporter-fehérje gyakran ugyanott fejeződik ki, mint ahol az ultrakonzervált szekvencia közelében levő gén. Vagyis, szól a cikkben is követett logika, ha ezek a szekvenciák hiányoznak, akkor annak valami szabályozási következménye kell legyen.
(Ezen a ponton, azt hiszem, megszellőztethetem azt a saját véleményt, hogy a fenti logika, bár nem alaptalan, koránt sem annyira alátámasztott, mint azt az elmélet manapság igen elterjedt elfogadottsága sugallná. A teljes igazsághoz tartozik ugyanis, hogy számos ultrakonzervált elem egyáltalán nem rendelkezik efajta enhancer aktivitással, ráadásul azt már tudjuk a valódi enhancerekben levő transzkripciósfaktor-kötőhelyekről, hogy azok ritkán maradnak nagyon konzerváltak, általában elég gyorsan változnak - mindez ismét az ellen szól, hogy az ultrakonzervált elemek "mezei" enhancerek lennének.)
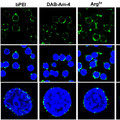
Végül 583 darab hCONDEL szekvenciát izoláltak, és ezek közül kettőt in vivo is megvizsgáltak, transzgénikus egerek segítségével.
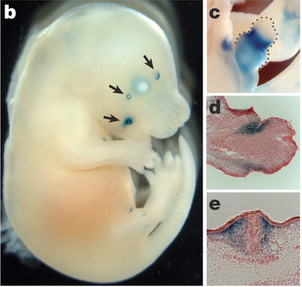 Az első az az androgén receptort (AR) kódoló gén közelében levő deléció volt. Amikor az emberből hiányzó, csimpánz szakaszt riporter-génhez kötötték, akkor utóbbi kifejeződését pl. a leendő bajusz és a pénisz területén lehetett megfigyelni.
Az első az az androgén receptort (AR) kódoló gén közelében levő deléció volt. Amikor az emberből hiányzó, csimpánz szakaszt riporter-génhez kötötték, akkor utóbbi kifejeződését pl. a leendő bajusz és a pénisz területén lehetett megfigyelni.
Az AR a tesztoszteron receptora, amelynek hatása egyébként dokumentált az egerek érzékszervként működő bajszának hosszára nézve. (A hatás nem drámai, de valós.) A mi történetünk szempontjából azonban nyilván az az érdekesebb, hogy a tesztoszteron egyszersmind a másodlagos nemi jellegek kialakulásáért is felelős, és a számos faj hímjeinek ivarszervét borító pénisztüskék tipikusan ilyenek.
A pénisztüskék (a poszt elején egy kandúr esetében jól megfigyelhetők) olyan kinövések a makk alatt, amelyeket kemény, keratin nevű anyag borít (de bennük semmi csontos elem nincs...), és általában két szerepet szoktak tulajdonítani nekik: érzékenyebbé teszik a hím ivarszervet, így a hím hamarabb képes a kopulációt befejezni, illetve egy előző hím által esetleg otthagyott kopulációs dugót is kitakarítják. (Ezeknek a dugóknak pont az a szerepük, hogy megnehezítsék a nőstény következő párzást, így időt nyerve, hogy a dugót elhelyező hím saját ivarsejtjeik célba érjenek.) Mindkét tulajdonság olyan fajokban előnyös, ahol erős a nőstényekért folytatott küzdelem, és az evolúciós verseny során számos érdekes, előnyt biztosító megoldás kialakult.
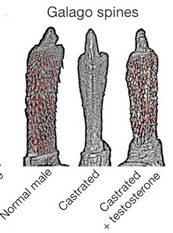 A pénisztüskék kialakulása szintén tesztoszteron szint függő: például kasztrált galágókban alig látszik belőlük valami, de tesztoszteron adagolásával mindez orvosolható.
A pénisztüskék kialakulása szintén tesztoszteron szint függő: például kasztrált galágókban alig látszik belőlük valami, de tesztoszteron adagolásával mindez orvosolható.
Embernél, ezt talán nem kell ragozni, nem találkozunk efajta keratinizált képletekkel, ami feltehetőleg a monogám kapcsolatokkal magyarázható. Hiányukban a párzás is tovább tart, amely intimitás akár segíthet is elmélyíteni a kapcsolatot. (Külön érdekesség, mindenképpen említést érdemel, hogy a szerzők az hCONDEL-riportergén konstrukciót emberi előbőrből készült sejtvonalakban is tesztelték, ahol az aktivitást mutatott, vagyis elvileg a képesség elvileg megvan a pénisztüskék képzésére emberben, csak épp az erre vonatkozó "utasítás" hiányzik.)
A magyarázat tehát az, hogy a párkapcsolatok változásával, őseink már nem tartottak igényt ezekre a képletekre. Így, ami elromolhat az el is romlik alapon, a szekvencia elvesződött. Apróbb problémák azért adódnak ezzel az interpretációval, de ezek boncolgatása előtt egy kis kitérő következik.
Ugyan a cikk-körüli médiafelhajtást elsősorban az AR-hez kapcsolódó hCONDEL keltette, legalább egy bekezdés erejéig érdemes a másik vizsgált génről is megemlékezni. Ez pedig a sejtciklust negatív módon szabályozó GADD45G előagy specifikus enhancerének tűnik. Ami azért fontos, mert ez az agyterület különösen megnövekedett bennünk, vagyis nem zárható ki, hogy egy ilyen szabályozóelem-deléció nyitotta meg a kaput az agyszövet kiterjedése előtt. (Általában hajlamosak vagyunk feltételezni, hogy ami bennünket különlegessé tesz, az új információ lehet a genomban, pedig, mint azt a MYH16 gén példáján át már bemutattam, van olyan, hogy már meglevő információ elvesztése tette lehetővé humán-specifikus változások megjelenését.)
Na, de visszatérve a nyilván minden olvasót izgalomban tartó pénisztüskékhez. Mi is a gond a szerzők okfejtésével. Az "A Primate of Modern Aspect" blog hosszú posztban járja körül, mennyire is állja meg a helyét az adatok interpretációja. Ugyanis a pénisz-tüskék száma és a monogámia közti összefüggés sem biztos, hogy statisztikailag szignifikáns, ráadásul a tüskék nagysága és a párzás-hossza közti korreláció nem annyira fix főemlősökben, mint azt a szöveg alapján gondolni lehetne, mert az orángutánokban, ahol lényegesen nagyobb méretűek a tüskék, sokkal tovább tart, mint csimpánzokban. Sőt, tulajdonképpen a csimpánzok esetében kérdéses, hogy beszélhetünk egyáltalán "tüskékről", amikor a libabőrnél alig nagyobb képletek vannak csak jelen, amelyek értelemszerűen alkalmatlanok bármifajta kopulációs dugó eltávolítására.
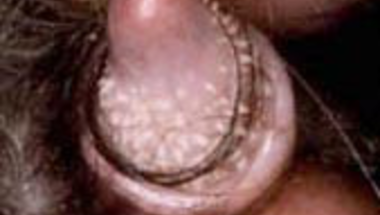
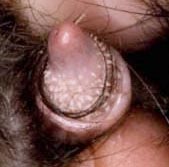
Ami még érdekesebb, hogy ilyen méretű képletek az emberben sem ritkák. A férfiak közel 30%-ában figyelhető meg a hímvessző papillomatosisa (18+ képek! - biztos, ami biztos ;-)), ami végső soron a tüskék csökevényes, atavisztikus megjelenését jelenti. Mivel méreteikben ezek a papillák összevethetőek a csimpánzok "tüskéivel", már rögtön nem is világos, hogy mi a hCONDEL szerepe.
Nagy kérdés persze, hogy ezekben a férfiakban is jelen van-e a szóbanforgó deléció? Mert ha igen, az azt jelentené, hogy a papillák/tüskék kialakulása mégsem kizárólag az ez által kódolt enhancernek köszönhető (sőt, elsősorban nem ennek), hanem léteznek más enhancerek is hasonló funkcióval, s az AR-hCONDEL lényegében olyan konzervált szekvencia, amely ugyan képes ilyen enhancer funkcióra is, de valódi szerepe valami más. Hogy az a "más" pontosan mi, azt majd jobban megértjük, ha végre világos képünk lesz az ultrakonzervált, nem kódoló szekvenciák biológiai szerepéről.
McLean, C., Reno, P., Pollen, A., Bassan, A., Capellini, T., et al. (2011) Human-specific loss of regulatory DNA and the evolution of human-specific traits. Nature 471: 216-219.