



Kevés dolgot tekintünk természetesebbnek, mint az ősz beköszöntésével bekövetkező levélsárgulást, majd -hullást. De vajon hányan gondoltunk bele, hogy mi is történik ilyenkor, mi az az élettani folyamat, ami szinte naptári pontossággal lezajlik milliárdnyi levélben minden ősszel? Mi kell ahhoz, hogy (Áprily szavaival élve) bükkök smaragd színét erezve fent az első pár vörös folt megjelenjen?
Mint minden összetettebb biológiai jelenségről, erről is természetesen akár könyvet lehetne írni, de azért most majd igyekszem itt ennél lényegesen rövidebben összefoglalni az egyes jelenségek hátterében álló folyamatokat.
Először is vegyük a színeket, amelyek az egyik legvarázslatosabb és legtöbb embert megihlető/lenyűgöző aspektusa az ősznek. Ha meg akarjuk érteni, hogy miért sárga/vörös színűek a lombhullató növények őszi levelei, akkor viszont először is annak kell utánajárnunk, hogy miért nem zöldek, mint mondjuk tavasszal, vagy nyáron.
A zöld színt a legismertebb növényi pigment, a levelekben levő klorofill adja, ami a növények fotoszintetizáló sejtorganellumának, a kloroplasztisznak a központi eleme. A klorofill által elnyelt fény fog aztán később a levelekben kémiai energiává alakulni, ami biztosítja a növény számos élettani folyamatát, köztük a szén-dioxid megkötését szerves vegyületek formájában. (A klorofill egyébként a kék- és vörös hullámhosszú fény elnyelésében jeleskedik, a zöld hullámhosszat szinte egyáltalán nem nyeli el, hanem visszaveri - ezért is látja, a visszavert fényt érzékelve, a mi szemünk zöldnek a klorofill tartalmú leveleket.)
A klorofill késő tavasszal és nyáron folyamatosan szintetizálódik a levél sejtjeiben, de ősz közeledtével a növény érzékeli a rövidülő nappalokat (erről egy kicsit később), aminek következtében a klorofill szintézise lassan leáll és különböző lebontó folyamatok elbontják a levelekben levő molekulákat, velük a korofillt is.
A lebontásra azért is szükség van, mert a télre készülve a növény igyekszik minden lehetséges tápanyagot visszanyerni a leveleiből, hogy utóbbiaktól később a lehető legkisebb áldozattal megszabadulhasson, ezzel is csökkentve a zord, szürke időjárásnak kitett felületét. A klorofill bomlástermékei már színtelenek, így a zöld szín fokozatosan elhalványul és ezáltal láthatóvá válnak a stabilabb sárga illetve narancssárga pigmentek (karotinoidok és flavonoidok), melyek korábban is jelenk voltak ugyan, de a klorofill dominanciája mellett nem voltak észrevehetőek. (A sárga/narancssárga színek élénksége egyéb jelenlevő vegyületektől is függ, például gingko esetében a sárga élénkségét a levelekben meglevő 6-hidorxikunerinsav teszi lehetővé.) Végül persze a karotinoidok és flavonoidok is lebomlanak, ekkor a levél elbarnul és sejtjei fokozatosan elhallnak (s végül persze lehull a fáról).
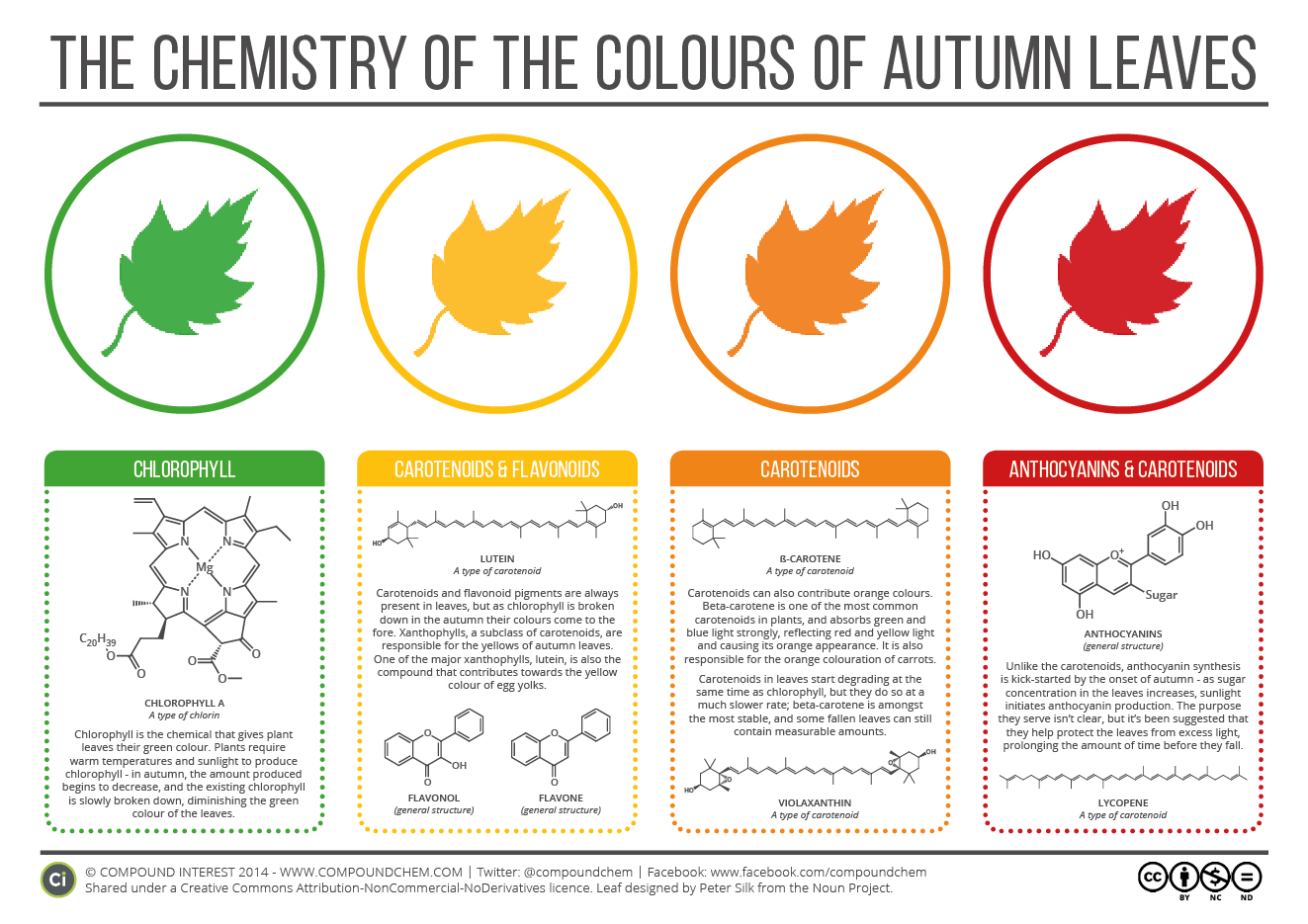
Mint a Compound Chemistry fenti ábrája is mutatja, az egyes levelekben megjelenő vörös szín egy kicsit más tészta: a szín forrását adó antocianin nevű pigmentek nem bomlástermékek, hanem ellenkezőleg, az ősz előrehaladtával a levelekben felgyülemlő cukrok és a napfény együttes hatása járul hozzá a szintézisükhöz. (Egyébként ugyanezen pigmentek a virágok vörös, lila, esetleg kékes színét is adják.) Mivel a növény ezeket aktívan termeli, egyértelmű, hogy valamilyen adaptív szerepük kell legyen, de nincs teljes konszenzus arról, hogy ez mi is lehet. A levélhullás körüli időszak különösen szenzitív időszak egy lombhullató növény életében, így abban a különböző elméletek egyezni látszanak, hogy az ideiglenesen megjelenő antocianinok védő szerepet látnak el. Csak míg egyesek szerint a fénytől óvják a klorofillját lebontó levelet, mások amellett kardosokodnak, hogy a növényevőket riasztja el ebben az időszakban.
Bármilyen átmeneti színezetet ölt is a levél a végső sora az elhullás (szaknyelven abszcisszió). Ez megint csak egy nagyon szabályozott folyamat, amely során a levél tövénél, az ún. abszcissziós zónában a növény felöli oldalon égy védőréteg alakul ki, majd felette meggyengül a sejtek közötti kapcsolat és a levél lehull.
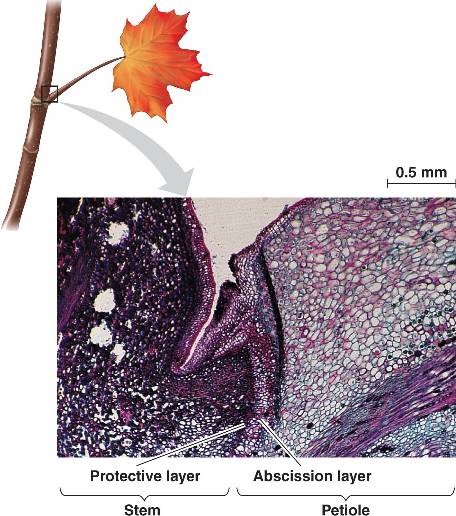
A folyamat ütemezése növényi hormonokon múlik, különösen fontos az auxin és az etilén. Előbbi folyamatosan termelődik a növekvő és működő levélben, és egyéb funkciói mellett biztosítja, hogy a levél tövénél levő sejtek ne figyeljenek az etilénre. Ősszel azonban a lebontó folyamatok túlsúlyba kerülésével párhuzamosan leáll az auxin szintézise, az etiléné pedig megnő és ez katalizálja egyes sejtfal-lebontó enzimek termelését az abszcissziós zóna környékén, a már előbb ismertetett eredménnyel.
De vajon miként érzi a növény, hogy ősz van? Komplex érzészervek, mint szem, fül, orr, híján, hogyan vehet tudomást a körülötte zajló folyamatokról? A válasz az, hogy nincs szükség annyira szofisztikált érzékszervekre, mint amilyenekkel mi rendelkezünk ahhoz, hogy egy élőlény tudatában legyen annak, milyen napszakban vagyunk, és épp rövidülnek-e a nappalok, vagy hosszabbodnak. Erre tökéletesen megfelelnek egyszerű molekuláris szenzorok, amilyenekkel már egysejtű élőlények is rendelkeznek, a növények meg pláne.
Az egyik ilyen fontos molekuláris szenzor a fitokróm nevű fehérje, amelynek kiindulási formájában (Pr) a nappali fényben levő vörös hullámhosszú komponens egy strukturális változást okoz, és az így keletkező ún. "aktív formája" (Pfr) a fehérjének mindenféle válaszreakciót katalizál. A Pfr azonban instabil, és alkonyatban gyakori távoli-vörös tartományú fényhullámok aktívan, a sötétben levés pedig passzívan visszaalakítja Pr-é. A két forma aránya ennek fényében (éérted, fényében...) jól tükrözi a világos és sötét periódusok hosszát, amit a növény arra tud használni, hogy az év múlását számontartsa.
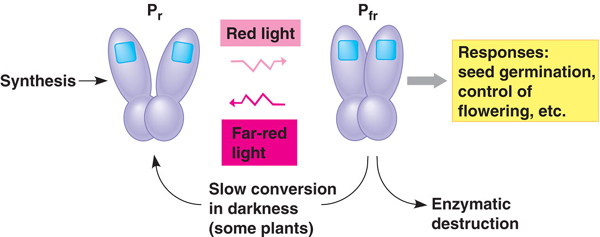
A fitokróm formák arányában történő változás épp ezért lehet fontos szabályozója a hideg téli időszakra való felkészülésnek is - na meg a virágozásra való felkészülésnek, de ez már tényleg egy másik történet.
Aki csak a levélsárgulás általános folyamatára volt kíváncsi az itt tulajdonképpen meg is állhat, az ínyenceknek azonban még álljon itt egy extra evolúciós csavar, amit nem is olyan rég írtak le. A levél sárgulása, vagyis tápanyagokban való elszegényedése ugyanis rossz hír azon élőlények számára, amelyek a zöld levelekkel táplálkoznának. És ne is a nagytestű növényevőkre gondoljunk most, hanem olyan kisméretű fajokra, mint az almalevél-aknázómoly lárvája. Táplálékának szezonalitása elég pontosan behatárolja, hogy ez a faj mikor tud maga is szaporodni, hiszen a sárguló-lehulló levél már nem képes elegendő táplálékot adni a fejlődő fiatal állatnak és az éhen pusztul.
Illetve úgy tűnik, van azért egy különleges megoldás, amivel az aknázómoly ki tudja tolni az egyébként elkerülhetetlent. Megfigyelték ugyanis, hogy a sárguló almafa-levelekben a molylárvák körül zöld "szigetek" maradtak fenn, amelyek tápanyagban és az örgedést gátló, citokininnek nevezett növényi hormonokban gazdagok, így ideális környezetet biztosítva a fejlődő állatnak. Magyarán az élősködő valamiképpen képes befolyásolni környezetét és az egyébként öregedő levélszövet hozzá közeli részeit olyan jelekkel ellátni, amelyek azokat megtévesztik.
A dolog mikéntje nem teljesen tisztázott, de erős jelek mutatnak abba az irányba, hogy ráadásul a jelenségért nem is közvetlenül maga a molylárva a felelős, hanem a benne található, kvázi-szimbionta baktérium, a Wolbachia.

A bakteriális kapcsolatnak a legegyszerűbb bizonyítéke, hogy antibiotikummal kezelt rovarok közelében a zöld szigetek képződése elmarad, bár ezek az anyagok csak prokariótákra hatnak közvetlenül, vagyis a moly sejtjeit direktben nem károsítják.
De hogy pontosan miként éri el közvetlen, vagy közvetve a Wolbachia a citokintermelés bekapcsolását az még nem tisztázott, mindenesetre ez csak egy újabb fantasztikus trükk ennek a mindenütt jelen levő, és rovarok tömegeit lényegében saját rabszolgájává tevő baktérium ingujjában.
Ougham HJ, Morris P, Thomas H (2005) The colors of autumn leaves as symptoms of cellular recycling and defenses against environmental stresses. Curr Top Dev Biol 66:135-60.
Kaiser W, Huguet E, Casas J, Commin C, Giron D (2010) Plant green-island phenotype induced by leaf-miners is mediated by bacterial symbionts. Proc Biol Sci 277(1692):2311-9.
Glover BJ, Martin C (2012) Anthocyanins. Curr Biol 22(5):R147-50.




