 Az evolúció által katalizált morfológiai változásoknak mindig fejlődéstani okai vannak, ugyanis a meglevő fejlődési program megváltoztatása/átírása lesz az elsődleges oka a kifejlett állatban megjelenő jelleg megjelenésének.
Az evolúció által katalizált morfológiai változásoknak mindig fejlődéstani okai vannak, ugyanis a meglevő fejlődési program megváltoztatása/átírása lesz az elsődleges oka a kifejlett állatban megjelenő jelleg megjelenésének.
A denevérek szárnyának kialakulásával már többször is foglalkoztunk itt a blogban, de míg az eddigi tanulmányok leginkább korábbi fejlődéstani ismereteink alapján keresték azokat a géneket, amelyeknek fontos szerepe lehet a szárny létrejöttében, addig a napokban megjelent tanulmány-páros, az újgenerációs szekvenálási technikáknak köszönhetően egy kicsit "elfogulatlanabb", előfeltételezéseket nélkülöző megközelítést alkalmazott.
Korábban már fény derült arra, hogy az úgynevezett BMP-jelátviteli útvonal különböző tagjai miként járulnak hozzá a kéz csontjainak megnyúlásához, illetve hogyan biztosítják az ujjpercek közti bőrred fennmaradását.
Most, a Nature Genetics-ben megjelent cikk egyrészt a denevér genom fehasználásával összeveti a mellső és hátsó végtag fejlődése során felfedezhető génexpressziós, ill. epigenetikai különbségeket, másrészt beazanosít olyan genomi régiókat, amelyek a denevérek evolúciója során különösen gyorsan változtak - s így feltehető, hogy közük lehet a csoport jellegzetes morfológiai újításának, a szárnynak a kialakításában. Ezek az ún. Bat Accelerated Regions (BAR) szekvenciák jelentik aztán egy PLOS Genetics cikk alapanyagát, és ezekkel fogunk mi is jobban foglalkozni.
A BAR szekvenciákat ugyanis viszonylag egyszerűen klónozni, majd egy könnyen detektálható riportergénhez csapva, egérembriókban megnézni, hogy van-e valamilyen enhanszer-funkciójuk (annak idején ezt a megközelítést használták erszényes farkas "gének feltámasztásához" is).
A kutatók összesen 166 BAR szekvenciát azonosítottak, amelyek többsége >20 kb távolságban volt a legközelebbi transzkripciós starthelytől. Harmincnyolc esetében olyan transzkripciós faktorok vannak a közelben, amelyeknek a végtagfejlődésben leírt szerepe már jó ideje ismert, ráadásul a BAR-ok előszeretettel (értsd véletlennél gyakoribban) helyezkednek el egymás közelségében.
Különböző megfontolások alaján öt BAR szekvenciát választottak ki a további vizsgálatokhoz és mint azt az alábbi ábra jól mutatja, ezek mindannyian végtagokban fejtenek ki génexpressziót (az ábrán a denevér BAR-ok és a megfelelő egérszekvenciák enhanszer-aktivitása közti különbséget látjuk).
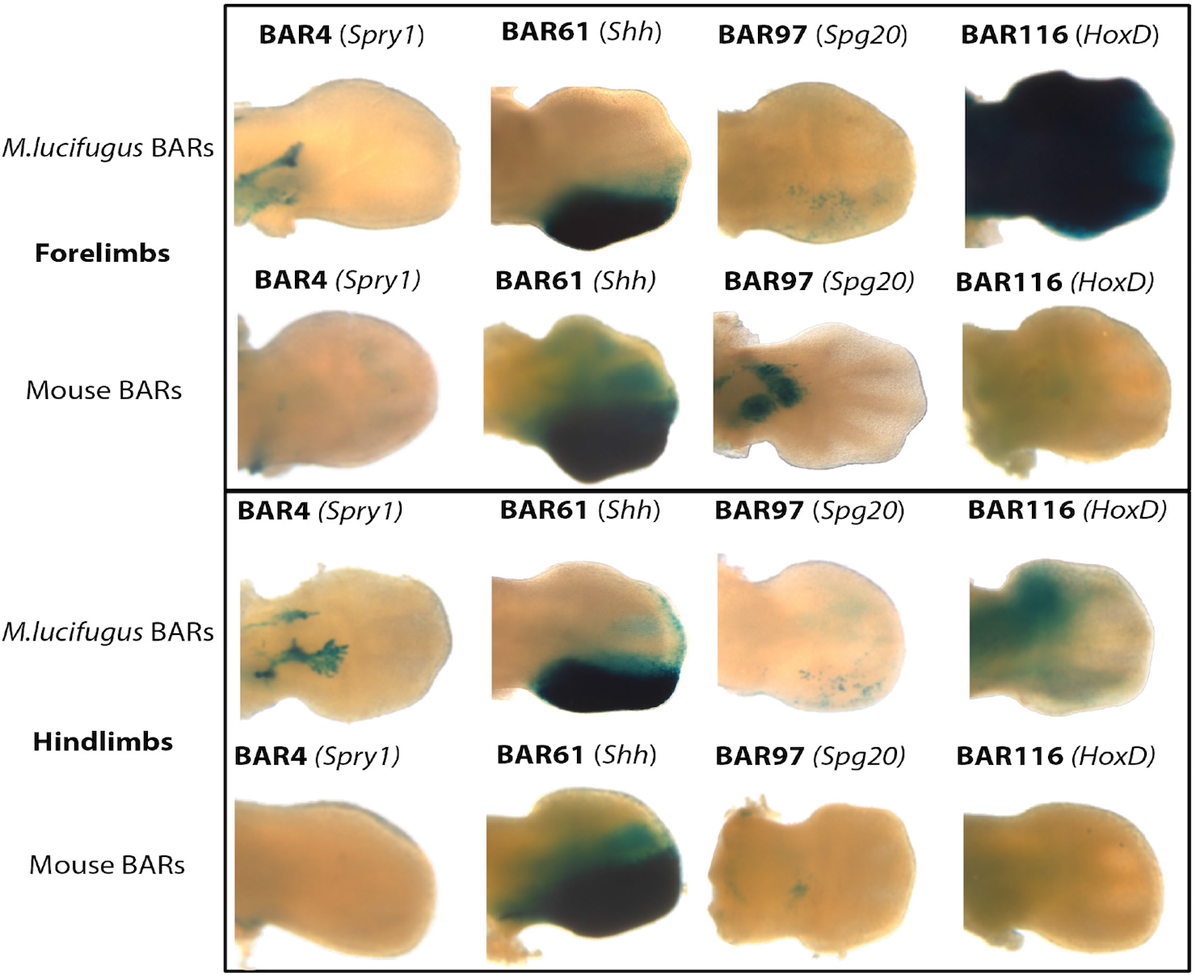
A vizsgáltakból különösen érdekesnek bizonyult a BAR116, ami a HoxD génklaszter melletti génsivatagban található. Mivel a HoxD gének végtagfejlődésben betöltött szerepe régóta ismert, ráadásul a BAR116 enhanszerként tényleg drámai változást okozott, különösen a mellső végtagban, a riportergén expressziójában, jó okkal lehetett feléttelezni, hogy érdemes megturkálni a denevérembriók HoxD expresszióját, mert ott is eltérések lesznek láthatók, az egérben tapasztalhattok képest.
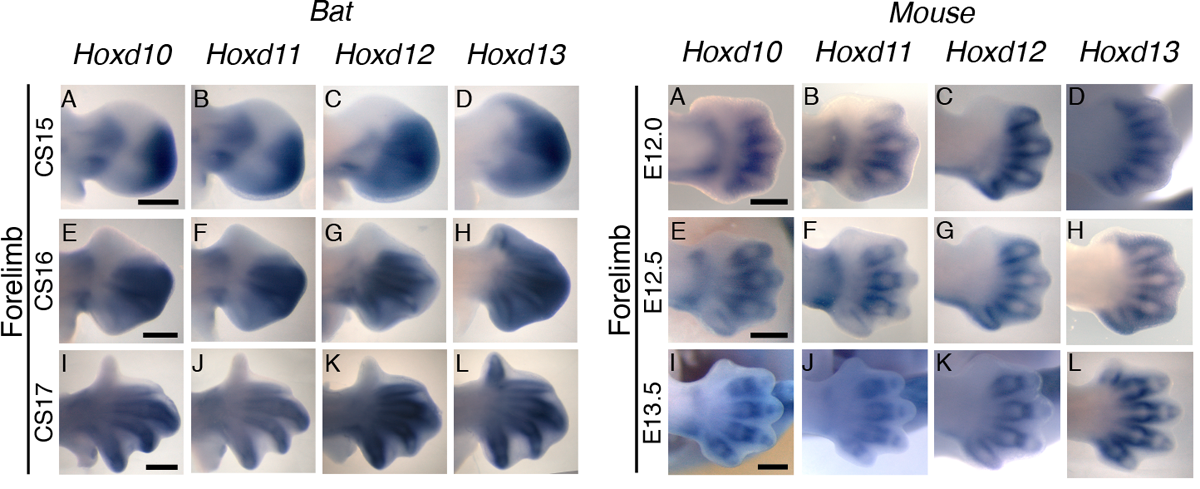
És valóban ezt tapasztalták, a végtagfejlődésben fontos HoxD gének, a megfelelő fejlődési stádiumban, általában intenzívebb expressziót mutattak denevérekben, mint egér megfelelőik egerekben.
Hogy persze ez konkrétabban mit jelent az egy nyitott kérdés. Minden felhajtás ellenére siralmasan keveset tudunk a HoxD faktorok target-génjeiről, így nem tudjuk megnézni, hogy mit jelent ez a megváltozott HoxD expresszió rájuk vonatkoztatva. Ahogy jelenleg még nem tudjuk azt sem megvizsgálni, hogy mi lenne, ha mondjuk a BAR116-t kiütnénk denevérembriókban. A technológia (a CRISPR-k révén) ezt lassan lehetővé teheti, de azért még valószínűleg jó-pár évet kell várni.
Booker BM, Friedrich T, Mason MK, VanderMeer JE, Zhao J, et al. (2016) Bat Accelerated Regions Identify a Bat Forelimb Specific Enhancer in the HoxD Locus. PLoS Genet 12(3):e1005738
Eckalbar WL, Schlebusch SA, Mason MK, Gill Z, Parker AV, et al. (2016) Transcriptomic and epigenomic characterization of the developing bat wing. Nat Genet doi: 10.1038/ng.3537.