Jó pár évvel ezelőtt, a blog hajnalán, az egyik legizgalmasabb és legfelkapottabb kérdés a biológiában az volt, hogy a különböző fajok (alfajok) morfológiai sajátosságai miképpen alakulnak ki a fejlődés során. Az evodevo tudománya - mert arról van szó - azóta kicsit kiment a mainstream divatból, de ha engem kérdezne bárki, szerintem továbbra is az egyik legizgalmasabb kérdés, amivel biológus foglalkozhat.
Jó pár évvel ezelőtt, a blog hajnalán, az egyik legizgalmasabb és legfelkapottabb kérdés a biológiában az volt, hogy a különböző fajok (alfajok) morfológiai sajátosságai miképpen alakulnak ki a fejlődés során. Az evodevo tudománya - mert arról van szó - azóta kicsit kiment a mainstream divatból, de ha engem kérdezne bárki, szerintem továbbra is az egyik legizgalmasabb kérdés, amivel biológus foglalkozhat.
A terület egyik úttörője, Sean B. Carroll mellett David Kingsley volt, aki azóta is azzal a nagyon egyszerű paradigmával foglalkozik, hogy milyen genetikai különbségek találhatók a háromtüskés pikó páncélos, tüskés tengeri és páncéltalan, tüskétlen édesvízi formái között.
Ahogy azt már korábban leírtam, ezzel a megközelítéssel leltek arra a pitx1 szabályozó mutációra, ami a tüskék eltűnését eredményezi, illetve így térképezték fel csontlemezek redukciójáért felelős ectodysplasin (eda) gént érintő mutációt is.
Most egy újabb mutációt határoztak meg, ami az édesvízi halakban a kevesebb csontlemez kialakulásáért, illetve a meglevő lemezek méretének megváltozásáért felel.
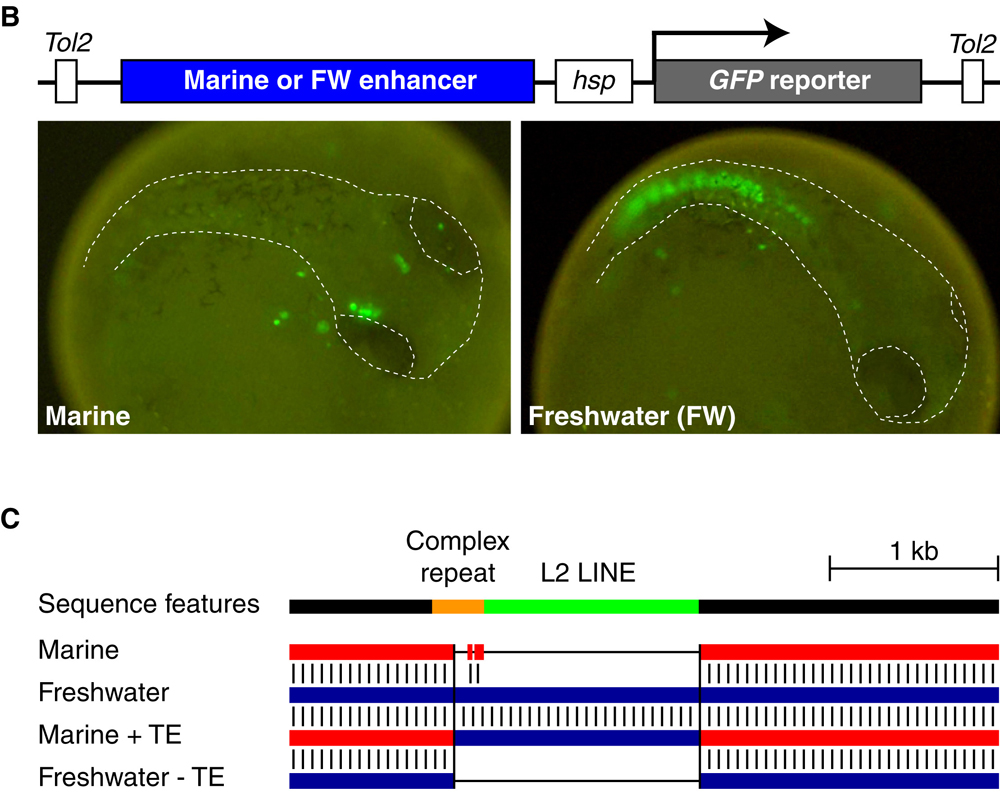 A klasszikus genetikai térképezés a gfd6 nevű gén közelében tette a mutációt, ami pont olyasmi, amit az ember találni szeretne: a Gdf6 fehérje ugyanis a BMP fehérjék csoportjába tartozik, amely rövidítés a Bone Morphogenetic Protein kifejezésből származik - azaz, már régóta tudjuk, hogy az ilyen típusú fehérjéknek szerepe van a csontok fejlődésében.
A klasszikus genetikai térképezés a gfd6 nevű gén közelében tette a mutációt, ami pont olyasmi, amit az ember találni szeretne: a Gdf6 fehérje ugyanis a BMP fehérjék csoportjába tartozik, amely rövidítés a Bone Morphogenetic Protein kifejezésből származik - azaz, már régóta tudjuk, hogy az ilyen típusú fehérjéknek szerepe van a csontok fejlődésében.
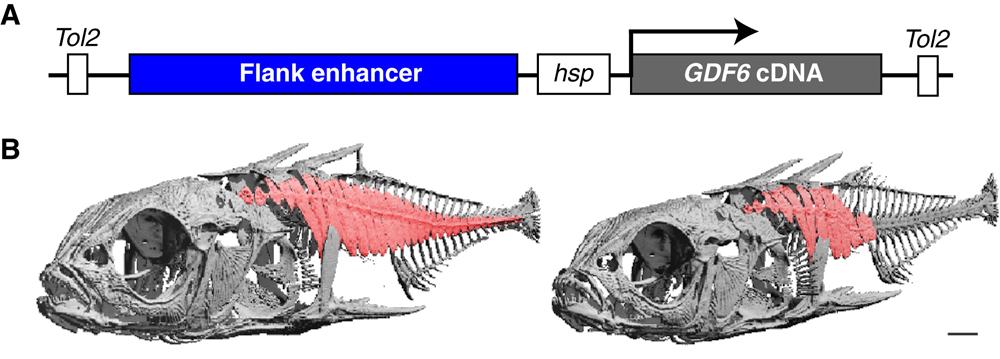
A mutáció megint egy szabályozó mutációnak bizonyult, ráadásul, ellentétben pl. a pitx1 esetével, amikor az édesvízi populációkban a gén egyik expressziós doménje elveszett, itt egy új expressziós domén megjelenésével állunk szemben: az édesvízi pikók megfelelő genomi régióját egy riportergén (pl. GFP) mögé téve a test teljes hosszában megfigyelhetünk expressziót a fejlődés egy korábbi szakaszában, olyan helyeken is, ahol a tengeri állatokban nem. Az oka ennek különleges (bár nem teljesen meglepő): egy ugrálóelem jelent meg az édesvízi pikók genomjában itt, és számos más példa alapján tudjuk, hogy ezek a mobilis genetikai elemek általában a génexpressziót elősegítő promótereket és enhancereket hurcolnak magukkal, amelyek elsősorban a saját genetikai anyaguk kifejezésére kellenek, de ha elég megengedő a genomi környezet, akkor a szomszédos génekre is hatással lehetnek.
Ami azonban a dolog legszebb bizonyítása, hogy ha ezzel az enhancerrel riportergén helyett GDF6-t hajtunk meg, és bevisszük tengeri fajokba, akkor nagyon hasonló elváltozást tapasztalunk, mint amit a tengeri -> édesvízi átmenet során az evolúció is produkált.
Épp ezért, kapcsolódva egy másik munkájukhoz, amely során olyan potenciális enhancereket keresnek, amelyek az emlősökben (de legalább főemlősökben) nagyon konzerváltak, viszont belőlünk hiányoznak, megnézték, hogy az emberi GDF6 gén környékén találunk-e ilyen szakaszt. A válasz röviden az, hogy találunk, és lehet, hogy ennek az eltűnése lényeges lépése lehetett a Homo génusz evolúciójának.
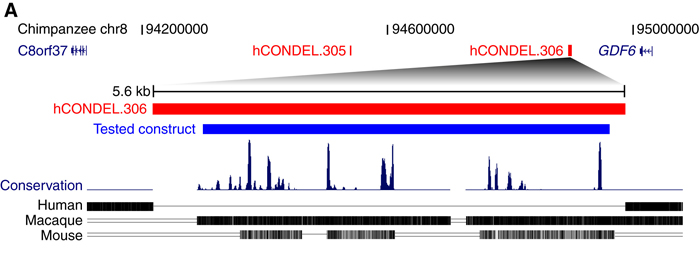
A szóbanforgó enhancer-jelölt (hCONDEL.306) ugyanis egy riportergénhez kapcsolva egerekben nagyon érdekes expressziós mintázatot mutatott: csak a test hátsó részében fejeződött ki, különösen erőteljesen a hátsó végtag ujjaiban. Ott mindenféle szövetben megtaláljuk, például a nagylábbujjak távolító izmában is, ami különösen felkeltette Kingsley-ék érdeklődését. Mégpedig azért, mert ez az izom az emberi evolúció során részben elsatnyult, hasonlóan bizonyos rágóizmokhoz. Utóbbiak esetében mára sikerült azonosítani a folyamat molekuláris hátterét (a MYH16 gén működésképtelenné válása) és valami hasonlót szerettek volna a GDF6 kapcsán Kingsley-ék elérni.
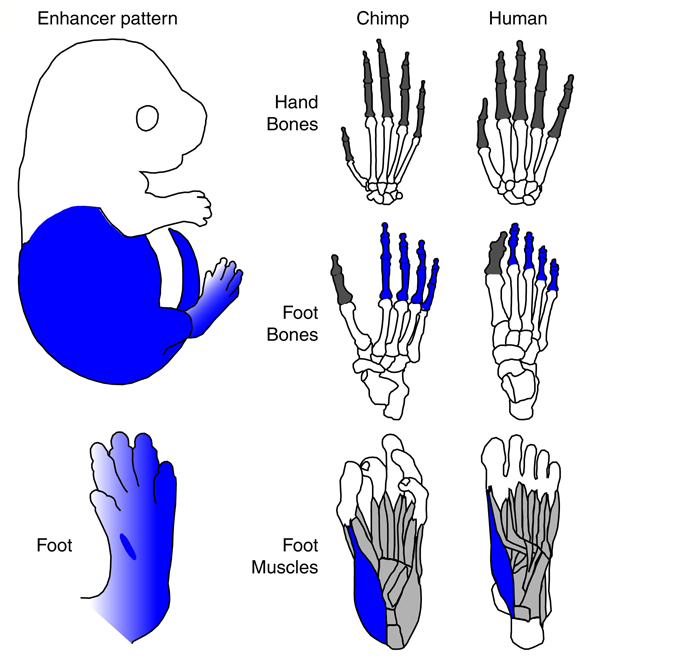
Mivel itt nem egy pszeudogénesedésről van szó, hanem egy feltételezett szabályozó mutációról, ez sokkal körülményesebb, és nem is sikerült nekik minden kétséget kizáróan (ehhez kb. csimpánzokban kellene kiütni a specifikus enhancert és nézni, mi történik - nem pont az a kísérlet, amit etikailag meg lehetne igazán okolni, és akkor még a technikai nehézségekről nem is szóltunk). De, azért az expresszión túl még egy fontos dolgot talált a csoport, amivel alá lehet támasztani az általuk favorizált evolúciós szcenáriót: azokban az egerekben, ahol kiütik a Gdf6 gént meg lehet figyelni, hogy a hátsó végtag ujjainak csontjai rövidebbek lesznek. Ez pedig megint egy olyan változás, amit az ember vs. csimpánz vonzatban is megtalálunk.
De ha elhisszük, ha nem a hCONDEL.306 történetét, azt aligha lehet elvitatni, hogy a gdf6 fontos szerepet játszik a tüskéspikók evolúciójában, hasonlóan a pitx1 és eda esetéhez. És már ez is pont elég lett volna, hogy foglalkozzunk vele :-).
Indjeian VB, Kingman GA, Jones FC, Guenther CA, Grimwood J, et al. (2016) Evolving New Skeletal Traits by cis-Regulatory Changes in Bone Morphogenetic Proteins. Cell doi: 10.1016/j.cell.2015.12.007.