 Amint az előző részből láttuk, a gombák elsődleges anyagcserefolyamatai az energiaszerzést (a sejtlégzés vagy az erjesztés révén) valamint a sejtalkotó makromolekulák szintézisét biztositják. Ezeken a reakciókon túl azonban léteznek bizonyos gombákban ún. másodlagos anyagcsereutak, amelyek termékei a másodlagos anyagcseretermékek. Ezek olyan kis molekulatömegű, biológiailag aktiv vegyületek, amik a gomba alapvető életfolyamataihoz ugyan nem nélkülözhetetlenek, de valamely, evolúciós szempontból előnyös tulajdonsággal vértezik fel az ezeket előállitó élőlényeket. Ezek az előnyök néha logikusan kikövetkeztethetők, ám néha homály fedi azt, hogy mire is jó az adott gombának egy bizonyos anyagot termelni. A legismertebb ilyen vegyületek talán az antibiotikumok, amik a konkurrens baktériumok elleni harc eszközei, de hogy a szintén mikroszkópkius gombák által termelt mérgező mikotoxinok vagy a különböző rákellenes, immunszupresszáns, virus- és gombaölő hatású vegyületek milyen előnyhöz juttatják az azokat termelő szervezeteket, sok esetben rejtély.
Amint az előző részből láttuk, a gombák elsődleges anyagcserefolyamatai az energiaszerzést (a sejtlégzés vagy az erjesztés révén) valamint a sejtalkotó makromolekulák szintézisét biztositják. Ezeken a reakciókon túl azonban léteznek bizonyos gombákban ún. másodlagos anyagcsereutak, amelyek termékei a másodlagos anyagcseretermékek. Ezek olyan kis molekulatömegű, biológiailag aktiv vegyületek, amik a gomba alapvető életfolyamataihoz ugyan nem nélkülözhetetlenek, de valamely, evolúciós szempontból előnyös tulajdonsággal vértezik fel az ezeket előállitó élőlényeket. Ezek az előnyök néha logikusan kikövetkeztethetők, ám néha homály fedi azt, hogy mire is jó az adott gombának egy bizonyos anyagot termelni. A legismertebb ilyen vegyületek talán az antibiotikumok, amik a konkurrens baktériumok elleni harc eszközei, de hogy a szintén mikroszkópkius gombák által termelt mérgező mikotoxinok vagy a különböző rákellenes, immunszupresszáns, virus- és gombaölő hatású vegyületek milyen előnyhöz juttatják az azokat termelő szervezeteket, sok esetben rejtély.
 A gombák összes rendszertani csoportja közül a penészgombák bioaktiv vegyületeit vizsgálták-vizsgálják a legaktivabban. 1922-ben Harold Raistrick, egy angol biokémikus a penészgombák anyagcseretermékeit kutatva több, mint 200 másodlagos metabolitot izolált és azonositott be. A penicillin alig hat évvel későbbi felfedézése után egymást hajszolva kerestek a kutatók hasonló antibakteriális anyagokat a penészgombák anyagcseretermékei között. Ma már több, mint tizezer gomba-eredetű másodlagos metabolit ismert, melyek kémiai szerkezete igencsak változatos. Érdekes módon, mig az elsődleges anyagcserében résztvevő enzimek génjei a gombák genomjában elszórtan találhatók meg, addig a másodlagos metabolitokat előállitó enzimeket kódoló gének szorosan egymás mellett, klaszterekbe tömörülve helyezkednek el. Ennek több oka is lehet, például e gének közös eredete (egy adott metabolitot csak egy szűk rendszertani csoport tagjai termelik), de akár az együttes szabályzásuk is. Legtöbbször ugyanis csak akkor aktiválónak ezek a másodlagos anyagcsereutak, amikor a gomba már megfelelő méretűre nőtt, és nekiáll spórákat termelni. Az ivartalan szaporodásra való átállást biztositó jelátviteli útról kiderült, hogy (többek között) ez is szabályozza a másodlagos anyagcserében szerepet játszó enzimeket.
A gombák összes rendszertani csoportja közül a penészgombák bioaktiv vegyületeit vizsgálták-vizsgálják a legaktivabban. 1922-ben Harold Raistrick, egy angol biokémikus a penészgombák anyagcseretermékeit kutatva több, mint 200 másodlagos metabolitot izolált és azonositott be. A penicillin alig hat évvel későbbi felfedézése után egymást hajszolva kerestek a kutatók hasonló antibakteriális anyagokat a penészgombák anyagcseretermékei között. Ma már több, mint tizezer gomba-eredetű másodlagos metabolit ismert, melyek kémiai szerkezete igencsak változatos. Érdekes módon, mig az elsődleges anyagcserében résztvevő enzimek génjei a gombák genomjában elszórtan találhatók meg, addig a másodlagos metabolitokat előállitó enzimeket kódoló gének szorosan egymás mellett, klaszterekbe tömörülve helyezkednek el. Ennek több oka is lehet, például e gének közös eredete (egy adott metabolitot csak egy szűk rendszertani csoport tagjai termelik), de akár az együttes szabályzásuk is. Legtöbbször ugyanis csak akkor aktiválónak ezek a másodlagos anyagcsereutak, amikor a gomba már megfelelő méretűre nőtt, és nekiáll spórákat termelni. Az ivartalan szaporodásra való átállást biztositó jelátviteli útról kiderült, hogy (többek között) ez is szabályozza a másodlagos anyagcserében szerepet játszó enzimeket.
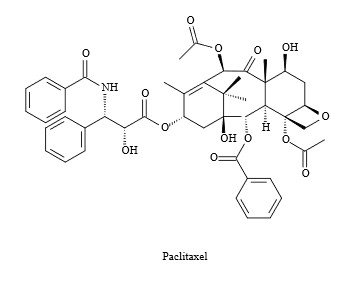 Mig az állatokra főként az elsődleges anyagcserefolyamatok jellemzőek, számos növény is képes másodlagos metabolitok előállitására, amely között van halálos méreg, vagy gyógyhatású anyag is. Ezek egy részéről kiderült, hogy a növény sejtjeiben élő endoszimbionta (a növénnyel szimbiózisban élő, vagyis nem kártevő vagy parazita) gombák is képesek termelni ugyanazokat az anyagokat, amit a növény. Ilyen például a prosztata-, mell-, tüdő- és petefészekrák terápiában használt taxol (paclitaxel). Erről sokáig úgy gondolták, hogy kizárólag a tiszafa (Taxus brevifolia) termeli, ám rajta kivül még a mogyoróban (Corylus avellana) is megtalálható, illetve e növények sejtjeiben élő szimbionta gombák is képesek a taxol szintézisére. Az amúgy bizonyos patogén gombák növekedését gátló (fungicid) anyag (amire a taxol-termelő endofita gombák természetszerűen rezisztensek) előállitásához szükséges enzimeket 13 gén kódolja a tiszafában. A taxoltermelő gombák, mint pl. a mogyoró-szimbionta Penicillium aurantiogriseum genomjának megszekvenálása alapján úgy tűnik, hogy a taxol bioszintézis nem a logikusnak tűnő laterális géntranszferrel került át a növényekből a gombákba, hanem a gombákban a növényekkel párhuzamosan, külön úton alakult ki.
Mig az állatokra főként az elsődleges anyagcserefolyamatok jellemzőek, számos növény is képes másodlagos metabolitok előállitására, amely között van halálos méreg, vagy gyógyhatású anyag is. Ezek egy részéről kiderült, hogy a növény sejtjeiben élő endoszimbionta (a növénnyel szimbiózisban élő, vagyis nem kártevő vagy parazita) gombák is képesek termelni ugyanazokat az anyagokat, amit a növény. Ilyen például a prosztata-, mell-, tüdő- és petefészekrák terápiában használt taxol (paclitaxel). Erről sokáig úgy gondolták, hogy kizárólag a tiszafa (Taxus brevifolia) termeli, ám rajta kivül még a mogyoróban (Corylus avellana) is megtalálható, illetve e növények sejtjeiben élő szimbionta gombák is képesek a taxol szintézisére. Az amúgy bizonyos patogén gombák növekedését gátló (fungicid) anyag (amire a taxol-termelő endofita gombák természetszerűen rezisztensek) előállitásához szükséges enzimeket 13 gén kódolja a tiszafában. A taxoltermelő gombák, mint pl. a mogyoró-szimbionta Penicillium aurantiogriseum genomjának megszekvenálása alapján úgy tűnik, hogy a taxol bioszintézis nem a logikusnak tűnő laterális géntranszferrel került át a növényekből a gombákba, hanem a gombákban a növényekkel párhuzamosan, külön úton alakult ki.
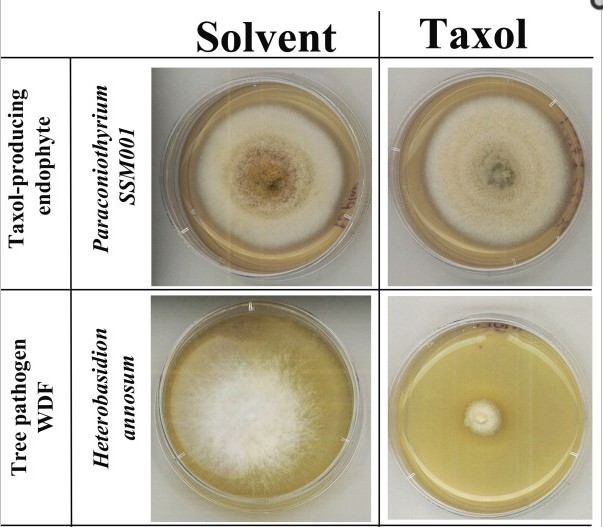
A taxol egyébként egy igencsak érdekes anyag. Évek óta ismert, hogy a mikrotubulusokhoz kapcsolódik, ezáltal gátolja a sejtosztódást. A legújabb, a rákbetegek szervezetében ténylegesen előforduló koncentrációt alkalmazó kisérletek eredménye alapján azonban az osztódás-gátló hatása koncentráció függő. A fiziológiás, kezelés alatti koncentrációja nem fagyasztja be az osztódásnak indult sejteket, hanem abnormális kromszóma-szétválást okoz, ezáltal eleve halálraitélt utódsejteket eredményez (lásd a lenti videót).
Calvo et al Relationship between Secondary Metabolism and Fungal Development, Microbiol. Mol. Biol. Rev. September 2002 vol. 66 no. 3 447-459
Keller NP et al Fungal secondary metabolism - from biochemistry to genomics. Nat Rev Microbiol. 2005 Dec;3(12):937-47.
Gunatilaka (2010). Fungal secondary metabolites. In AccessScience. McGraw-Hill Education. Retrieved from http://accessscience.com/content/fungal-secondary-metabolites/YB100063
Zasadil et al, Cytotoxicity of Paclitaxel in Breast Cancer Is due to Chromosome Missegregation on Multipolar Spindles Sci Transl Med 26 March 2014: Vol. 6, Issue 229, p. 229ra43
Soliman, S. S. M., C. P. Trobacher, R. Tsao, J. S. Greenwood and M. N. Raizada (2013). "A fungal endophyte induces transcription of genes encoding a redundant fungicide pathway in its host plant." BMC Plant Biol 13: 93.