Még valamikor a nyári uborkaszezon közepén futott egy kört a magyar médiában (is) a hír, hogy Olaszországban egy zebra apától és szamár anyától származó kis jövevény látta meg a napvilágot.
Még valamikor a nyári uborkaszezon közepén futott egy kört a magyar médiában (is) a hír, hogy Olaszországban egy zebra apától és szamár anyától származó kis jövevény látta meg a napvilágot.
Ippo, a kis zebramár, mint minden fajok közötti (interspecifikus) hibrid természetesen biológia érdekesség, de mint azt sok helyen megjegyezték sajnos aligha jelenti egy keverékfaj létrejöttét. Ugyanis akárcsak az öszvérek, vagy más hasonló "korcs" élőlények, Ippo minden valószínűség szerint steril.
Az ok amit a legtöbb helyen megadnak, az a szüleinek eltérő kormoszómaszáma: a zebrák 22, a szamarak pedig 31 pár kromoszómával rendelkeznek, ami alapján a zebramár zigóta sejtjei összesen 53 kromoszómát fognak hordozni. Ami, szól az érvelés, páratlan szám, így meióziskor életképtelen ivarsejtek alakulnak majd ki. (A lovakban 32 pár kromoszóma van, így az öszvérek testi sejtjei 63 kormoszómát hordoznak.)
A gond persze nem is annyira a páratlan kormoszómaszám, hanem inkább az, hogy ez önmagában olyan kromoszómaszerkezet-eltéréseket kódol, ami miatt az ivarsejtek jelentős része használhatatlan lesz.
Normális esetben az ivarsejtek képződési folyamata, a meiózis során (ha valaki már nem emlékszik, miként is zajlik az, itt felfrissítheti a tudását) a homológ kromoszómadarabok egymás mellé helyeződnek, lezajlódik az átkereszteződés (crossing-over) és az utódsejtekbe összességében egyenlő mennyiségű DNS kerül, normális kormoszómákon. Az eltérő kormoszómaszám azonban garantáljam hogy az átkereszteződések bizonyos része hibás legyen, részben mert a kromoszómák közepe, a kritikus centromér régió, más relatív helyzetbe kerül egyes génekhez képest, illetve a centromérákhoz képest a gének relatív sorrendje gyakran megváltozik (a miértet lásd alább).
Ha jól meghatározható géneket jelző, különleges kromoszómafestékekkel megfestjük a zebra, szamár és ló egyes kromoszómáit, akkor láthatni fogjuk, hogy az egyes vizsgált génpárok kromoszómán belüli (vagyis telomérákhoz és centromeronhoz viszonyított) sorrendje nagyon is változó lehet.
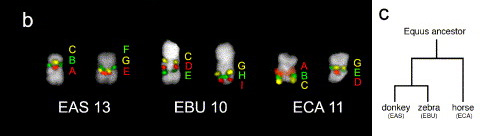
Márpedig, mint erre fentebb utaltam, ez azt jelenti, hogy az említett átkereszteződések aligha zajlódhatnak le minden gubanc nélkül.
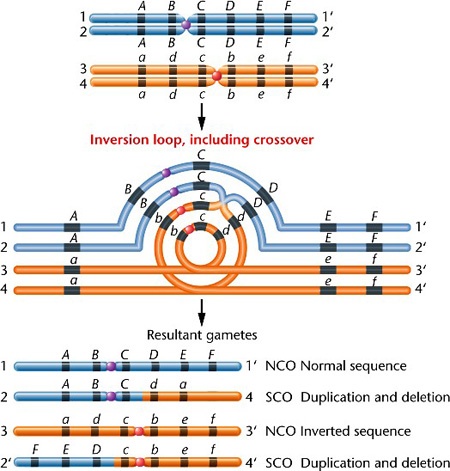 Ugyanis, ha az egyik kromoszómán inverziók vannak jelen, akkor a crossing-over után jó eséllyel olyan kromoszómák jönnek létre ahol teljes genomi régiók hiányoznak, mások pedig megduplázódva vannak jelen (hogy miért is, lásd a bal oldali ábrán). Ez pedig halálos kombináció, hiszen az utódsejtben így bizonyos géntermékek alig lesznek jelen, mások pedig sokkal nagyobb dózisban, mint az szükséges lenne. (Az itt mutatott, ún. pericentrikus inverziónak van egy paracentrikus párja, ami hasonló bonyodalmakat okoz, lásd itt.)
Ugyanis, ha az egyik kromoszómán inverziók vannak jelen, akkor a crossing-over után jó eséllyel olyan kromoszómák jönnek létre ahol teljes genomi régiók hiányoznak, mások pedig megduplázódva vannak jelen (hogy miért is, lásd a bal oldali ábrán). Ez pedig halálos kombináció, hiszen az utódsejtben így bizonyos géntermékek alig lesznek jelen, mások pedig sokkal nagyobb dózisban, mint az szükséges lenne. (Az itt mutatott, ún. pericentrikus inverziónak van egy paracentrikus párja, ami hasonló bonyodalmakat okoz, lásd itt.)
Az inverziók mellett az új centromérák kialakulása (ami velejárója a kromoszóma-szám növekedésnek), szintén bonyolíthatja a crossing-overek lezajlását, mert így, akárcsak a fent linkelt paracentrikus inverziók esetében könnyen két centromérrel renelkező dicentrikus, vagy centromérek nélküli acentrikus kromoszómák alakulnak ki. Előbbiek sejtosztódás során, amikor a két centromért az osztódási orsók más irányba húzzák, random helyen széttörnek, míg utóbbiak, centromér hiányában, láthatatlanok marandak a sejtosztódás mechanizmusa számára, így random sodródnak majd vagy egyik, vagy másik utódsejtbe.
Mivel ha a fajok közti távolság már akkor, hogy az kormoszómaszám különbségben is megnyilvánul, akkor ott jó eséllyel sok kromoszómaátrendeződés is jelen lesz, így az utódok nagyon valószínűleg elsöprő többségében életképtelen ivarsejteket fognak létrehozni, így sterilek lesznek.
Természetesen, a mellékelt ábrán látható hibás átkereszteződéshez még csak nem is kell eltérő kromoszómaszám, ugyanazon faj, azonos kromoszómaszámú két egyede közt is bekövetkezhet - ez a magyarázata annak, hogy sajnálatos módon vannak olyan személyek, akiknek sosem lehet saját gyereke, mert az ivarsejtjeik életképtelen genetikaianyag-kombinációt hordoznak.
Sőt, ha már ennyire belementünk a dolgokba, akkor az is említést érdemel, hogy még efajta kormoszómaátrendeződés hiányában is előfordul a sterilitás: pl. távoli rokon populációk egyedei közt létrehozott hibridek esetében. Mert, míg egy-egy populációban fontos fehérjekomplexeket alkotó fehérjék általában úgy koevolválódnak, hogy együttműködésük zavartalan legyen, különböző populációkban nem feltétlenül lesz azonos az evolúció iránya. S így, ha egy hibrid egyedben összekerülnek, már korántsem lesz olajozott a fehérjék együttműködése, sőt.
Érdekes megfigyelés, hogy a hímeket méginkább érint a sterilitás (vagyis ha nagynéha van termékeny öszvér, az nőstény lesz, tuti). Ez az ún. Haldane szabály - és több magyarázat is van arra, mi lehet a dolog hátterében. Az egyik, hogy az őket erősebben érintő szexuális szelekció miatt a hímek szexdeterminációjáért felelős gének evolúciója valamivel gyorsabb. Ennek következtében viszont a fent említett genetikai konfliktusok hamarabb is jelentkeznek esetükben.
(Az inverziós crossing-over kép forrása ez.)
Carbone L, Nergadze SG, Magnani E, Misceo D, Francesca Cardone M, et al. Evolutionary movement of centromeres in horse, donkey, and zebra. Genomics 87(6): 777-82.