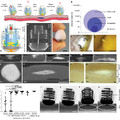
A béka hátsó lábában, a gekkók melső végtagjában és a mi kezünkben a számtalan homológ csontocska mellett is feltünő egy, fontos, közös vonás: mindannyiunknak öt ujja van. Az öt ujjúság a szárazföldi négylábú gerincesek nagy közös vonása, olyannyira, hogy egy tudományos kifejezés is születtet a sztereotip négylábú végtag leírására: pentadactyl végtag.
A béka hátsó lábában, a gekkók melső végtagjában és a mi kezünkben a számtalan homológ csontocska mellett is feltünő egy, fontos, közös vonás: mindannyiunknak öt ujja van. Az öt ujjúság a szárazföldi négylábú gerincesek nagy közös vonása, olyannyira, hogy egy tudományos kifejezés is születtet a sztereotip négylábú végtag leírására: pentadactyl végtag.
Az ettől való eltérés mindig különös és az evolúciósan csak egy irányba történik: a békák melső lábán, a madarak szárnyában, vagy a lovak patájában egyaránt kevesebb ujj figyelhető meg (négy, három, illetve egy). Az ötnél több ujjúság (az ún. polydactylia) kóros esetekben észlelhető, például a végtagok fejlődése apropóján korábban már emlegetett Sonic-hedgehog (Shh) gén mutációja okozhat ilyen elváltozást.
De vajon miért pont öt a normális, illetve lehette-e valamikor hat, vagy hét? Esetleg tizenkettő?
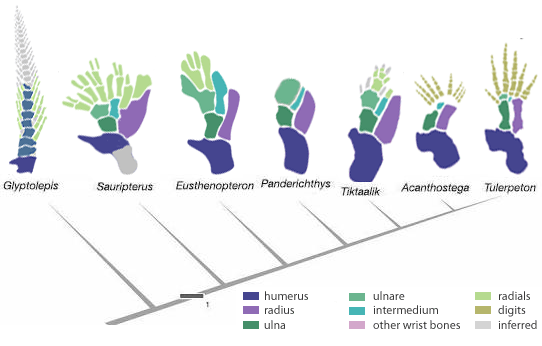
Ha a fosszilis anyagot tekintjük, akkor láthatjuk, hogy az öt nem is annyira evidencia. A négylábúak (Tetrapoda) evolúciójának hajnalán megjelenő Acanthostega illetve Tulerpeton végtagján egyaránt több ujjal rendelkezett (nyolccal illetve hattal - lásd alábbi ábrán), erősítve a gyanút, hogy a szelekció több lehetőséget végigzongorázott, mígnem a pentadactylia rögzült valamelyik távoli ősünkben. Hogy az ötnek van-e/volt-e evolúciós előnye pl. a hathoz képest, vagy rögzülése egyszerű evolúciós véletlen, arról igazából fogalmunk sincs, ugyanakkor érdekes annak a megértése, hogy miképp is szabályozódik az ujjak száma a fejlődés során, mert ez részben elárul valamit az evolúciójukról is.
Ha a végtagfejlődést tekintjük, akkor tudható, hogy a végtag disztális (testfaltól távolabbi) végének, vagyis a kéz(láb)fejnek a kialakulásáért egy bonyolult genetikai hálózat a felelős, amiben Shh mellett szerepet játszanak a Fibroblast growth factor (Fgf) gének, valamit a HoxA és HoxD kluszterek leghátsó génpárosai. A Shh egyben az egzes ujjak identitásáért is felelős, hiszen pl. a kis- és mutatóujj közti kinézetbeli (morfológiai) különbség annak a számlájára (is) írható, hogy az egyes ujj-kezdemények mennyi ideig vannak a géntermék hatásának kitéve.
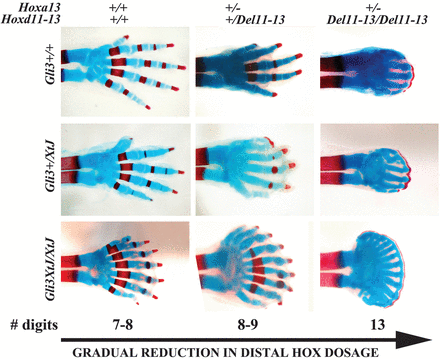 Ezért is meglepő talán, hogy ha a Shh-jelátvitelt teljesen gátoljuk (pl. a jelátviteli útvonal effektorát kódoló Gli3 gén kiütésével - Gli3XtJ) nem kevesebb ujjal találkozunk, hanem többel, egész pontosan hét-nyolccal. (A bal oldali ábrán embrionális egér végtagok porc-csont preparátumait lehet látni, ahol a porcot kék, a csontot pedig piros jelöli.)
Ezért is meglepő talán, hogy ha a Shh-jelátvitelt teljesen gátoljuk (pl. a jelátviteli útvonal effektorát kódoló Gli3 gén kiütésével - Gli3XtJ) nem kevesebb ujjal találkozunk, hanem többel, egész pontosan hét-nyolccal. (A bal oldali ábrán embrionális egér végtagok porc-csont preparátumait lehet látni, ahol a porcot kék, a csontot pedig piros jelöli.)
Az ujjak száma tovább növelhető, akár tizenháromig is, ha a már emlegetett Hox géneket is kiütjük (ami szintén nem triviális, hiszen ezek (is) az ujjidentitás kialakításában is szerepet játszanak). Mivel a Shh szerepe pont a Hox gének szabályozása, a jelek arra mutatnak, hogy a mintázatképző folyamat valahogy a Hox-aktivitástól függ.
De mi is lehet ez a mintázatképző folyamat? Hogy ezt jobban megértsük, vissza kell mennünk a fejlődési program egy korábbi szakaszába, amikor a végtagbimbóban kialakulnak a leendő porcosodási gócok, magyarán az a mintázat, ami a leendő ujjakat jelöli.
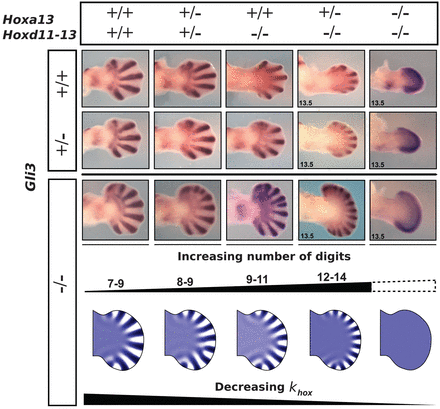 Ezeket a "proto"ujjakat a Sox9 gén kifejeződésének láthatóvá tételével mutathatjuk ki, a festődés eredményeként a lapát alakú végtagprimordiumon egy sugár irányú csíkokból álló mintázat jelenik meg. (A leendő ujjak közti terület sorsa egy másik történet.) A hangsúly a mintázaton van, ugyanis innen akár már fel is merülhet gyanúként, hogy esetleg egy Turing-mintázattal állunk szemben.
Ezeket a "proto"ujjakat a Sox9 gén kifejeződésének láthatóvá tételével mutathatjuk ki, a festődés eredményeként a lapát alakú végtagprimordiumon egy sugár irányú csíkokból álló mintázat jelenik meg. (A leendő ujjak közti terület sorsa egy másik történet.) A hangsúly a mintázaton van, ugyanis innen akár már fel is merülhet gyanúként, hogy esetleg egy Turing-mintázattal állunk szemben.
S valóban: mint kiderült tényleg lehet olyan reakció-diffúzió mechanizmust modellezni, amivel szimulálható, hogy a Hox-aktivitás függvényében, hogyan változhat dinamikusan az ujjak száma. Mindehhez ráadásul nem is kell az aktivátor és inhibitor molekulák pontos mibenlétét sem ismerni. Elég feltételezni, hogy minél erősebb a Hox- (illetve Fgf-) aktivitás, annál erőteljesebben gátlódik az inhibitor. Ezzel már elérhető, hogy míg a vad típusúhoz hasonlóan magas Hox/Fgf szint esetében viszonylag ritkásan helyezkednek el a vastag ujjak, addig a redukált Hox szinttel rendelkező mutánsokban sűrűbben fognak a vékony protoujjak feltűnni (és minél kisebb a Hox szint, annál inkább igaz ez).
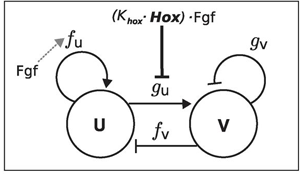 Eközben a Hox/Fgf aktivitás azt is meghatározza, hogy mekkora legyen az ujjak által a végtagbimbóban elfoglalt terület. Csökkenésükkor így az is megfigyelhető, hogy az ujjak egyre rövidebbek, a szélsőséges esetekben pedig ki sem alakulnak.
Eközben a Hox/Fgf aktivitás azt is meghatározza, hogy mekkora legyen az ujjak által a végtagbimbóban elfoglalt terület. Csökkenésükkor így az is megfigyelhető, hogy az ujjak egyre rövidebbek, a szélsőséges esetekben pedig ki sem alakulnak.
A Turing-modellünknek még kell egy sajátos csavarja legyen: ha a lapát-alakú végtagbimbót nézzük, akkor a sugarak közti távolság egyre nagyobb, ahogy az ujjhegyek felé tartunk. Márpedig, ha a mintázat periódusa a "kéztőnél" x, akkor az "ujjbegyek" magasságában már akár több ujjnak is lehetne hely. Vagyis kell egy olyan paraméter is, ami biztosítja, hogy a periódus az fokozatosan növekszik, a kézfej vége felé. Ezt ismét az Fgf-jelátvitel moduláló szerepében találhatjuk meg (hiszen ez amúgy is egy gradiens formájában van jelen, ahol a kéz végén a legnagyobb a ligandum koncentrációja, távolodva pedig fokozatosan csökken), ami az aktivátor szintézisét befolyásolhatja (Fgf -> f(u)).
Hát ez lehet az a nem túl egyszerű, de mégis elegáns mintázat-képző rendszer, amelynek az evolúciója végső soron létrehozta a pentadactyliát. Időgép hiányában nehezen megmondható, hogy az Acanthostega esetében mi eredményezte a nagyobb ujjszámot, így maradunk az ötletelésnél: vagy más paraméterekkel működött a mechanizmus, így nyolc ujj jöhetett létre, vagy ugyanezen paraméterek egy szélesebb végtagbimbó végén több ujjat tudtak létrehozni. Persze, ha szerencsénk van, egy nap valamilyen hal-mutánsok erről is valami érdekeset árulnak majd el.
(A végtagevolúciót mutató ábra a Devonian Times posztja alapján készült. Ha valaki belelkesült a csont-porc prepiktől, itt egy részletes protokoll, hogyan is csinálják őket.)
Sheth R, Marcon L, Bastida MF, Junco M, Quintana L, Dahn R, Kmita M, Sharpe J, Ros MA (2012) Hox genes regulate digit patterning by controlling the wavelength of a Turing-type mechanism. Science 338: 1476-80.