 Amikor egyszer régen írtam egy hosszú, bevezető posztot az emlős végtagok fejlődéséről, akkor az volt a nem is igen titkolt szándékom, hogy az egész amolyan felvezetésként szolgál majd a mellső végtag evolúcióját tárgyaló posztokhoz. Aztán teltek az évek és igazából csak nem sikerült időt szakítani a témára, pedig mindig lett volna anyag. Múlt hónapban azonban két olyan fontos cikk is kijött, ami miatt most mégis össze kell kapnom magam, nem is egy, hanem két poszt erejéig, mert ezekről hallgatni már-már bűn lenne. :-)
Amikor egyszer régen írtam egy hosszú, bevezető posztot az emlős végtagok fejlődéséről, akkor az volt a nem is igen titkolt szándékom, hogy az egész amolyan felvezetésként szolgál majd a mellső végtag evolúcióját tárgyaló posztokhoz. Aztán teltek az évek és igazából csak nem sikerült időt szakítani a témára, pedig mindig lett volna anyag. Múlt hónapban azonban két olyan fontos cikk is kijött, ami miatt most mégis össze kell kapnom magam, nem is egy, hanem két poszt erejéig, mert ezekről hallgatni már-már bűn lenne. :-)
Az első cikk azzal foglalkozik, hogy miképpen befolyásol(hat)ta az elnyújtott Hoxd13 expresszió a végtagok fejlődését. De, hogy ne rontsunk ajtóstól a házba, előbb lássuk, hogyan kerül elő egyáltalán ez a gén és egyáltalán, miben is különbözik egy egér és egy hal végtagja.
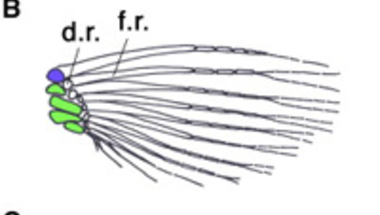
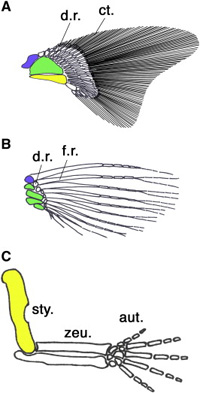
Ha az ősi gerinces végtagot vesszük szemügyre, ami ma is megfigyelhető egyes procoshalakban (lásd cápák - A), akkor azt látjuk, hogy a végtag ízesülését a vállövhöz három nagy csont biztosítja, ezek a propterygium (kék), mesopterygium (zöld) és metapterygium (sárga). Ezekhez apróbb csontok kapcsolódnak (distal radials - d.r.), amelyek az úszósugarakat tartják. Csontoshalak esetében (B) ez a szabásminta úgy egyszerűsödött, hogy a metapterygium elvesződött, és az úszósugarakat tartó apróbb csontok száma is redukálódott. A szárazföldi gerincesekben (C) viszont pont a metapterygium maradt egyedül meg (felkarcsontként), a disztális radiálisokból pedig kialakultak az alkarunk csontjai és a kéztőcsontok. (Némi nemű vita van arról, hogy az ujjak maguk az úszósugarak homológjai-e, vagy sem, de a jelenleg dominánsabbnak tűnő nézet szerint igen.)
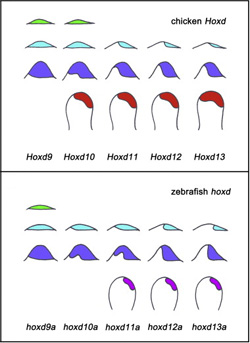 És akkor, hogy miként is jönnek mindehez a Hox gének. Mint arról a már említett bevezető posztban írtam, a Hox géneknek két fontos szerepe is van a tipikus emlős végtag fejlődésében: előbb, kollineárisan kifejeződve a kar csontok identitását határozzák meg, majd a végtagbimbó tetejében egy második hullámban az ujjak kialakulását segítik elő (és a Hoxd13 fejeződik ki a legszélesebb területen).
És akkor, hogy miként is jönnek mindehez a Hox gének. Mint arról a már említett bevezető posztban írtam, a Hox géneknek két fontos szerepe is van a tipikus emlős végtag fejlődésében: előbb, kollineárisan kifejeződve a kar csontok identitását határozzák meg, majd a végtagbimbó tetejében egy második hullámban az ujjak kialakulását segítik elő (és a Hoxd13 fejeződik ki a legszélesebb területen).
Sokáig úgy tűnt, hogy ez a második hullám csak a szárazföldi gerincesekre jellemző, de aztán előbb az ősi testfelépítést mutató halfajokról, majd a modern csontoshalakról (pl. zebradánió) is bebizonyították, hogy egy második hullámnak megfeleltethető génexpresszió is detektálható a fejlődés során. A különbség az, hogy ennek intenzitása és kiterjedtsége sokkal kisebb, mint a négylábúak (Tetrapoda) csoportjában.
A kérdés tehát adott: okozhat-e lényeges különbséget, ha mondjuk egy hal mellúszójában hossz(abb)an fejeztetjük ki a Hoxd13-t, megváltozik-e a morfológiája az úszónak, netán "végtagszerűbb" lesz-e?
Ennek megválaszolására egy spanyol kutatócsoport olyan transzgénikus halakat hoztak létre, ahol szabályozni lehetett, hogy a hoxd13 egy extra kópiája mikor kapcsoljon be (hoxd13a-GR). Ha ezt a "bekapcsolást a mellúszók kialakulásanak idejére időzítették, akkor jellegzetes elváltozásokat tudtak megfigyelni a fejlődő végtagokban.
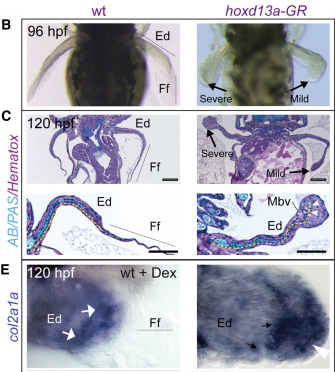 Az úszók megvastagodtak, a végükön furcsa kis bumpszlik jelentek meg. Amikor csont/porc specifikus festékekkel (pl. alciánkék - AB) festeték meg őket, azt találták, hogy az addig ezektől mentes úszóredő (finfold - Ff) is tartalmaz elcsontosodó sejteket. Sőt, azok a gének, amelyek korábban csak az úszó proximális részén fejeződtek ki, hirtelen megjelentek az úszóredő területén is.
Az úszók megvastagodtak, a végükön furcsa kis bumpszlik jelentek meg. Amikor csont/porc specifikus festékekkel (pl. alciánkék - AB) festeték meg őket, azt találták, hogy az addig ezektől mentes úszóredő (finfold - Ff) is tartalmaz elcsontosodó sejteket. Sőt, azok a gének, amelyek korábban csak az úszó proximális részén fejeződtek ki, hirtelen megjelentek az úszóredő területén is.
Minden arra utalt tehát, hogy alapvetően megváltozik a végtag fejlődése pusztán a hoxd13a külön-expresszió hatására.
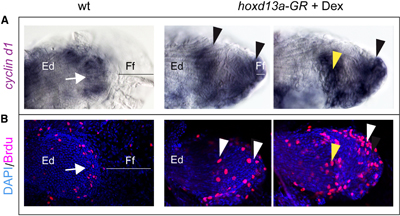
A bumpszlikban levő több sejt azt is engedte sejtetni, hogy ezekben az állatokban nagyobb méretű sejtproliferáció zajlott le a végtag-kezdeményekben.
És valóban ez a helyzet: osztódó sejteket jelölő festékekkel (BrdU), és a sejtciklusban fontos CcnD1 fehérje mRNS-ének a megfestésével egyaránt az látszott, hogy az egykori úszóredő sejtjei most már nem csendesednek el, nem lépnek ki az osztódási mókuskerékből, hanem nagyon is aktívan hoznak létre újabb és újabb sejteket.
 Az igazi csattanó persze az lenne, ha ezen a ponton azt lehetne mondani, hogy "és az így kezelt lárvákból felnövő kifejlett halakban már-már kézszerű struktúrák jelentek meg", de azért ennyire nem tökéletes a sztori. A kezelt lárvák sajnos meghalnak, mégpedig pont azért, mert az eldeformálódott úszó nem teszi lehetővé, hogy elég hatékonyan táplálkozzanak.
Az igazi csattanó persze az lenne, ha ezen a ponton azt lehetne mondani, hogy "és az így kezelt lárvákból felnövő kifejlett halakban már-már kézszerű struktúrák jelentek meg", de azért ennyire nem tökéletes a sztori. A kezelt lárvák sajnos meghalnak, mégpedig pont azért, mert az eldeformálódott úszó nem teszi lehetővé, hogy elég hatékonyan táplálkozzanak.
Vagyis két következtetést vonhatunk le: egyrészt, hogy a hoxd13 megnyilvánulásának időbeni és térbeni szabályozása, pontosabban annak változásai, fontos feltételei lehettek a komplexebb végtagok fejlődésének. Ugyanakkor az is nyilvánvaló, hogy mindezek a változások nem egyik pillanatról a másikra következtek be, hanem hosszú idő alatt, amiközben egyéb változások biztosíthatták, hogy a Tiktaalik és tsai. felé mutató új generációk is fittek maradjanak és sikeresen vadásszák le zsákmányaikat.
Ahn D, Ho RK (2008) Tri-phasic expression of posterior Hox genes during development of pectoral fins in zebrafish: implications for the evolution of vertebrate paired appendages. Dev Biol 322(1): 220-33.
Schneider I, Shubin NH (2012) Making limbs from fins. Dev Cell 23(6): 1121-2.
Freitas R, Gómez-Marín C, Wilson JM, Casares F, Gómez-Skarmeta JL (2012) Hoxd13 contribution to the evolution of vertebrate appendages. Dev Cell 23(6): 1219-29.