Méltatlanul elhanyagoltuk egyik kedvenc témánkat, a pigmentáció evolúcióját, de most remek apropó nyílik arra, hogy visszatérjünk rá.
Méltatlanul elhanyagoltuk egyik kedvenc témánkat, a pigmentáció evolúcióját, de most remek apropó nyílik arra, hogy visszatérjünk rá.
Az utóbbi időben egyre aktívabb Hopi Hoekstra csoportja most az egerek után egy másik kistermetű gerinces csoportot célzott meg: az Új Mexikóban élő gyíkokat.
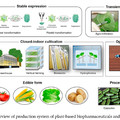
Egész pontosan három fajt, a keleti sövényleguánt (Sceloporus undulatus), a csíkos teju (Aspidoscelis inornata) és a Holbrookia maculata-t (magyar nevéért még nyomozok) tüntették ki figyelmükkel, mivel mindhárom fajnak ismertek "albínó" populációi, amelyek a Chihuahuan sivatag White Sands nevű gipszdűnéin élnek. Ebben a környezetben érelemszerűen előnyös fehérnek lenni, hiszen így kevésbé keltik fel az állatok a ragadozóik figyelmét, s mivel a táj kb. 6000 éve alakult ki, így ez egy viszonylag friss formáját jelenti az evolúciónak.
Az fő gyanúsított (mint ilyen esetekben lenni szokott) a pigmentációs útvonal kulcsgénje, a melanocortin-1-receptor (mc1r) volt: tudjuk, hogy ha ez a receptor-gén olyan mutációt szed össze, ami folytán folyamatosan aktív lesz, az fekete mintázathoz, ún. melanizációhoz vezet, míg a működését tönkretevő mutációk, világos (vöröses) színkomplexiót okoznak. (Itt egy kicsit szakmai, de zanzásított képi összefoglaló az útvonal működéséről.)
Méltatlanul elhanyagoltuk egyik kedvenc témánkat, a pigmentáció evolúcióját, de most remek apropó nyílik arra, hogy visszatérjünk rá.
Az utóbbi időben egyre aktívabb Hopi Hoekstra csoportja most az egerek után egy másik kistermetű gerinces csoportot célzott meg: az Új Mexikóban élő gyíkokat.
Egész pontosan három fajt, a keleti sövényleguánt (Sceloporus undulatus), a csíkos teju (Aspidoscelis inornata) és a Holbrookia maculata-t (magyar nevéért még nyomozok) tüntették ki figyelmükkel, mivel mindhárom fajnak ismertek "albínó" populációi, amelyek a Chihuahuan sivatag White Sands nevű gipszdűnéin élnek. Ebben a környezetben érelemszerűen előnyös fehérnek lenni, hiszen így kevésbé keltik fel az állatok a ragadozóik figyelmét, s mivel a táj kb. 6000 éve alakult ki, így ez egy viszonylag friss formáját jelenti az evolúciónak.
Az fő gyanúsított (mint ilyen esetekben lenni szokott) a pigmentációs útvonal kulcsgénje, a melanocortin-1-receptor (mc1r) volt: tudjuk, hogy ha ez a receptor-gén olyan mutációt szed össze, ami folytán folyamatosan aktív lesz, az fekete mintázathoz, ún. melanizációhoz vezet, míg a működését tönkretevő mutációk, világos (vöröses) színkomplexiót okoznak. (Itt egy kicsit szakmai, de zanzásított képi összefoglaló az útvonal működéséről.)
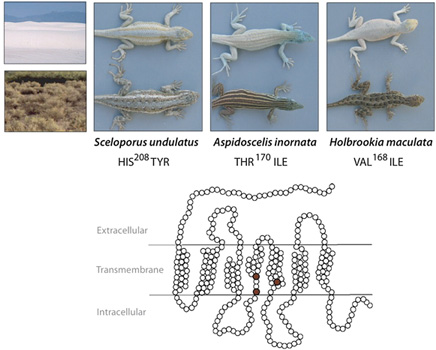 A három esetből kettőben be is bizonyosodott, hogy valóban az Mc1r működésével van gond és ez az oka (legalábbis részben) az észlelt "albinizmusnak". (Az ábrán a receptor vázlatába az egyes fajok esetében előforduló jellegzetes mutációk lettek feltüntetve. Ezek pontos mibenléte a fajok képe alatt található.) A két külön fajnál, azonban teljesen más módon került sor a receptor működésének modulálására. (A Holbrookia sp. esetében valahol máshol keresendő a fehér szín oka - hiába hordoz az mc1r génjük is egy jellegzetes mutációt, annak, úgy tűnik, nincs hatása.)
A három esetből kettőben be is bizonyosodott, hogy valóban az Mc1r működésével van gond és ez az oka (legalábbis részben) az észlelt "albinizmusnak". (Az ábrán a receptor vázlatába az egyes fajok esetében előforduló jellegzetes mutációk lettek feltüntetve. Ezek pontos mibenléte a fajok képe alatt található.) A két külön fajnál, azonban teljesen más módon került sor a receptor működésének modulálására. (A Holbrookia sp. esetében valahol máshol keresendő a fehér szín oka - hiába hordoz az mc1r génjük is egy jellegzetes mutációt, annak, úgy tűnik, nincs hatása.)
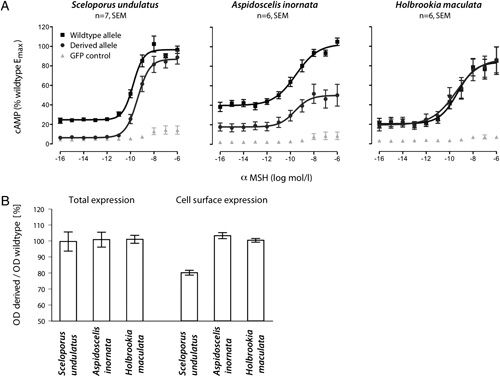
Annyi közös, mind a sövényleguánok, mind a tejuk esetében, hogy a mutáns receptorok jelenlétében a sejt sokkal kisebb válaszreakciót generál, mint azt tenné hasonló körülmények között, nem mutáns ("vad") receptor jelenlétében (az alábbi ábrán ezt méri a felszabaduló cAMP). Azonban míg a tejuk esetében ennek az az oka, hogy a receptor kevésbé "vezeti a jelet", a sövényleguánoknál a jelek szerint arról van szó, hogy a mutáns receptor alacsonyabb valószínűséggel kerül ki a sejt felszínére (és kevesebb receptor értelemszerűen kisebb jelet generál).
A mutációk típusa azért sem mindegy, mert ez határozza meg, hogy hatásuk domináns vagy recesszív. Mivel az Mc1r molekulák a sejt belsejében dimerizálódnak és csak ezt követően kerülnek ki a membránba, látható, hogy heterozigóta sövényleguáok esetében, hiába van jelen a "vad" allél is, azt a mutáns allél visszatartja a citoplazmába, így aztán a hatása ilyen körülmények közt is érezteti magát. Magyarán, a mutáció domináns. A tejuk esetében viszont a "vad" allél, ha jelen van, kompenzálni tudja a mutáns allél működőképtelenségét, azaz ez egy recesszív mutáció, aminek hatása csak akkor lesz látható, ha egy egyed kizárólag a mutáns mc1r allélokat hordozza.
Mivel a szelekció a kinézeten, vagyis a fenotípusokon "dolgozik" mindez kihatással van a különböző élő populációk genetikai összetételére is. A leguánok esetében az "albínó" populációkban is jelen lesz a vad típusú allél, hiszen heterozigóta formában az még mindig fehér színt eredményez, viszont a hagyományos környezetben élőkből teljesen hiányozni fog a mutáns allél (mert nem épp adaptív a fűcsomók közt fehéren rohangálni). A tejuk esetében pedig épp fordítva: a "klasszikus" populációkban is megtalálható a mutáns allél, ellenben a vadtípusú variáns csak nyomokban van jelen a dűnelakók génállományában.
Összességében a példa szépen mutatja, hogy a konvergens evolúció sem mindig ugyanazt az útvonalat járja be, még ha gyakorta ugyanazon az útvonalon keresztül érezteti a hatását.
(Az első kép innen való.)
Rosenblum EB, Römpler H, Schöneberg T, Hoekstra HE. (2010) Molecular and functional basis of phenotypic convergence in white lizards at White Sands. PNAS 107(5): 2113-2117.