Ami elromolhat, az el is romlik, tartja Murphy örökbecsűje, és természetesen ez igaz az élővilágra vonatkoztatva is. Azzal a kitétellel megtoldva, hogy a szelekció szigorából csak olyan dolog szabadulhat ki (vagyis romolhat el, a fitnessz vészes lecsökkenése/megszünése nélkül), amit az élőlény már nem, vagy nem nagyon használ.
Ami elromolhat, az el is romlik, tartja Murphy örökbecsűje, és természetesen ez igaz az élővilágra vonatkoztatva is. Azzal a kitétellel megtoldva, hogy a szelekció szigorából csak olyan dolog szabadulhat ki (vagyis romolhat el, a fitnessz vészes lecsökkenése/megszünése nélkül), amit az élőlény már nem, vagy nem nagyon használ.
Ilyenek például természetesen örök kedvenceink, a csökevényszervek, vagy ezek DNS szintű analógjai, a pszeudogének.
Amikor utóbbiakról egy átfogó poszt keretében szót ejtettem anno, már említettem, hogy a legszebb példák egyike a szaglóreceptorokhoz (olfactory receptors - OR) tartozik.
Ami elromolhat, az el is romlik, tartja Murphy örökbecsűje, és természetesen ez igaz az élővilágra vonatkoztatva is. Azzal a kitétellel megtoldva, hogy a szelekció szigorából csak olyan dolog szabadulhat ki (vagyis romolhat el, a fitnessz vészes lecsökkenése/megszünése nélkül), amit az élőlény már nem, vagy nem nagyon használ.
Ilyenek például természetesen örök kedvenceink, a csökevényszervek, vagy ezek DNS szintű analógjai, a pszeudogének.
Amikor utóbbiakról egy átfogó poszt keretében szót ejtettem anno, már említettem, hogy a legszebb példák egyike a szaglóreceptorokhoz (olfactory receptors - OR) tartozik.
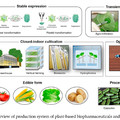
 Az emberben, és tágabb rokonságában, az óvilági majmokban, ez a receptorcsalád durván pszeudogenizálódott (vagyis számos OR gén, bár felismerhető a szekvenciában, de nem működőképes). Ez azért különösen feltűnő, mert a másik nagy "majomcsaládban", az újvilági majmokban (ill. az emlősökben általában) az OR géneknek igencsak fontos szerepük van és a szelekciós nyomás nem engedi, hogy mutációkat halmozzanak fel. Az egyetlen, de mint mindjárt látni fogjuk, instruktív kivétel a bőgőmajom (howler monkey), amely esetében szintén megugrott az OR pszeudogének száma.
Az emberben, és tágabb rokonságában, az óvilági majmokban, ez a receptorcsalád durván pszeudogenizálódott (vagyis számos OR gén, bár felismerhető a szekvenciában, de nem működőképes). Ez azért különösen feltűnő, mert a másik nagy "majomcsaládban", az újvilági majmokban (ill. az emlősökben általában) az OR géneknek igencsak fontos szerepük van és a szelekciós nyomás nem engedi, hogy mutációkat halmozzanak fel. Az egyetlen, de mint mindjárt látni fogjuk, instruktív kivétel a bőgőmajom (howler monkey), amely esetében szintén megugrott az OR pszeudogének száma.
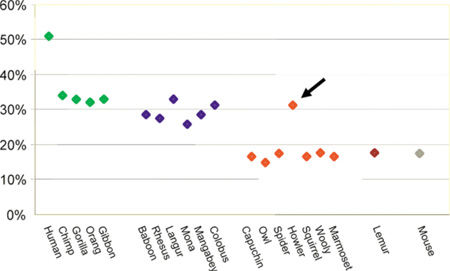
 Mi a közös a bőgőmajmokban és az óvilági majmokban, ill. hogyan különböznek ezek a többi újvilági majomtól? A kulcsjelleg ezúttal valószínűleg a színlátás fejlődése. A mellékelt törzsfán piros vonal vezet mindazokhoz a fajokhoz, amelyek három, különböző hullámhosszra érzékeny opszin gént hordoznak, így trikromatikus színekben látják a világot (az emlősök esetében az alapállapot a két opszin gén).
Mi a közös a bőgőmajmokban és az óvilági majmokban, ill. hogyan különböznek ezek a többi újvilági majomtól? A kulcsjelleg ezúttal valószínűleg a színlátás fejlődése. A mellékelt törzsfán piros vonal vezet mindazokhoz a fajokhoz, amelyek három, különböző hullámhosszra érzékeny opszin gént hordoznak, így trikromatikus színekben látják a világot (az emlősök esetében az alapállapot a két opszin gén).
Ennek a jellegnek az adaptív funkcióját akkor érthetjük meg jobban, ha egy kicsit belegondolunk egy átlagos majomfaj táplálkozási szokásába: a menü jelentős részét gyümölcsök teszik ki. Egy gyümölcs érettségi fokát pedig vagy szagán vagy színén keresztül lehet megítélni. Így azokban a fajokban, amelyekben az egyik opszin gén duplikációja révén kialakulhatott a trikromatikus látás, már nem volt annyira létfontosságú a kifinomult szaglás, így az OR gének jelentős százaléka fokozatosan mutálódni kezdett, s végül működésképtelenné vált. (Az újvilági bőgőmajmok saját csoportjukban egyedülálló nonkonformizmusa mind a színlátás, mind a szagreceptorok pszeudogenizálódása terén különösen szemléletessé teszi a példát.)
A színlátás minőségbeli ugrása persze csak az egyik potenciális út az OR gének nélkülözhetősége válása felé. Egy másik az olyan közegbe való átszokás, ahol eleve nincs szükség egy széles OR-repertoárt kihasználni.
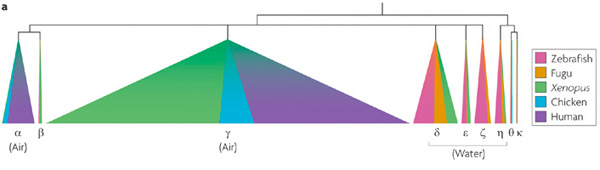
A fenti ábra az OR gének családfáját szemlélteti, azt ábrázolva, hogy az egyes OR gén-alcsaládok tagjai milyen fajokban jelentek meg először. (Ez az "először"persze relatív, itt azt jelenti konkrétan, hogy a rendelkezésre álló mely genomokban találjuk meg őket, és ebből következtetünk az evolúciós eredetükre.) Jól látható, hogy a szárazföld meghódítása a négylábú gerincesek által a gének számában hatalmas ugrást jelentett, aminek az a racionális magyarázata, hogy sokkal több szagmolekula tud a levegőben "közlekedni", mint a vízben. Megfordítva a logikát, ennek alapján azt gondolhatjuk, hogy az olyan fajokban, amelyek "visszaköltöztek" a vízbe, hirtelen sok szaglóreceptor fölösleges lett (hiszen a közeg miatt sosem találkozhatott az állat azokkal a szagmolekulákkal, amelyek az adott OR-t aktiválják), és hát mint már említettem, ami elromolhat, az jó eséllyel el is romlik...
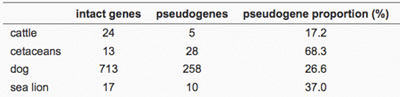 A két adódó példa közül az egyik a ceteké. Egy átlagos cet OR génjeinek közel hetven százaléka pszeudogén, míg a szegről-végről rokon szervasmarháknál ez az arány húsz százalék van.
A két adódó példa közül az egyik a ceteké. Egy átlagos cet OR génjeinek közel hetven százaléka pszeudogén, míg a szegről-végről rokon szervasmarháknál ez az arány húsz százalék van.
A ragadozó tengeri oroszlánok negyven százalékos pszeudogén aránya ennél azonban jóval szerényebb, különösen, ha a kutyák 26.6%-hoz hasonlítjuk (bár a kutyák esetében ez a szám bő nagyságrenddel több aktív receptort takar), aminek az a valószínű magyarázata, hogy bár ezek az állatok a vízben vadásznak, de életük jelentős részét (az utódnevelésre való különös tekintettel) a szárazon töltik, így számukra nem indifferensek a levegőben terjedő szagok.
A vízi életmód és az OR pszeudogének közti összefüggésre adódó másik példa a tengeri kígyók esete.
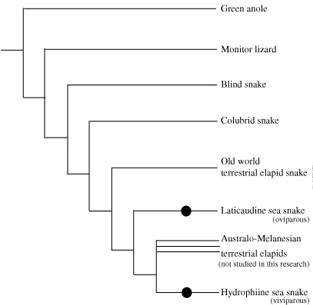 Ahogy a cetek, ezek az fajok is szárazföldi ősökből alakultak ki. Jelenleg két bővebben vett csoportba oszthatók: az egyikbe azok a fajok tartoznak, amelyek egész életüket a vízben töltik le, és elevenszülők (vagyis még szaporodáshoz sem kell a partra kúszniuk) - ezeket a szaknyelv vivpar fajoknak nevezi -, a másikba azok tartoznak akik életük jelentős részét a tengerben töltik, de azért tojással szaporodnak és tojás rakáshoz bizony kimennek a partra (ovipar fajok).
Ahogy a cetek, ezek az fajok is szárazföldi ősökből alakultak ki. Jelenleg két bővebben vett csoportba oszthatók: az egyikbe azok a fajok tartoznak, amelyek egész életüket a vízben töltik le, és elevenszülők (vagyis még szaporodáshoz sem kell a partra kúszniuk) - ezeket a szaknyelv vivpar fajoknak nevezi -, a másikba azok tartoznak akik életük jelentős részét a tengerben töltik, de azért tojással szaporodnak és tojás rakáshoz bizony kimennek a partra (ovipar fajok).
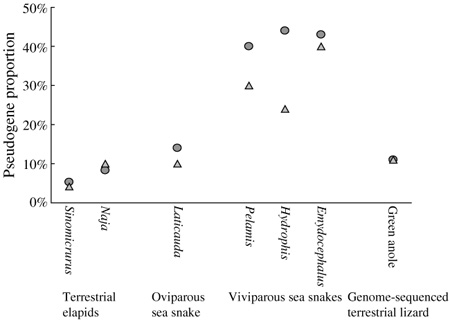
Az életmódokból adódó analógia a két csoport ill. a cetek meg a tengeri oroszlánok közt így jól láthatóan dobja magát, és a hasonlóság az OR pszeudogének arányára is kiterjed: a vivipar tengeri kígyó fajok szaglóreceptor génjeinek kb. 40%-a működésképtelen, míg az ovipar fajok esetében ez az arány alig több, mint a szárazföldi fajok szűk tíz százaléka (az alábbi ábrán a körök a teljes OR génállományra vonatkoznak, a háromszögek, pedig a Colubroidae szupercsaládra jellemző OR gén-alcsalád nélkül).
Gilad Y, Przeworski M, Lancet D. (2004) Loss of olfactory receptor genes coincides with the acquisition of full trichromatic vision in primates. PLoS Biol 2(1): E5. doi: 10.1371/journal.pbio.0020005
Kishida T, Kubota S, Shirayama Y, Fukami H. (2007) The olfactory receptor gene repertoires in secondary-adapted marine vertebrates: evidence for reduction of the functional proportions in cetaceans. Biol Lett 3(4): 428-30. doi: 10.1098/rsbl.2007.0191
Kishida T, Hikida T. (2009) Degeneration patterns of the olfactory receptor genes in sea snakes. J Evol Biol doi: 10.1111/j.1420-9101.2009.01899.x