 Új gének keletkezésének egyik jól bevált módja a génduplikáció, amikor a szóbanforgó DNS szakasza a genomnak valamilyen úton-módon, a sejtosztódás során megkétszereződik. Az így létrejövő extra másolat aztán viszonylag szabadon mutálódhat (hiszen az eredeti funkciót ellátja a másik kópia), ami leggyakrabban ahhoz vezet, hogy a gén működésképtelenné válik, de néha az is előfordul, hogy valamilyen új funkcióra tesz szert, ami ha előnyös az élőlény számára, jó eséllyel rögzülni fog (ezt nevezik a szakzsargonban "neofunkcionalizációnak").
Új gének keletkezésének egyik jól bevált módja a génduplikáció, amikor a szóbanforgó DNS szakasza a genomnak valamilyen úton-módon, a sejtosztódás során megkétszereződik. Az így létrejövő extra másolat aztán viszonylag szabadon mutálódhat (hiszen az eredeti funkciót ellátja a másik kópia), ami leggyakrabban ahhoz vezet, hogy a gén működésképtelenné válik, de néha az is előfordul, hogy valamilyen új funkcióra tesz szert, ami ha előnyös az élőlény számára, jó eséllyel rögzülni fog (ezt nevezik a szakzsargonban "neofunkcionalizációnak").
Egy harmadik, a köztudatban kevésbé forgó lehetőség (bár egyáltalán nem biztos, hogy ritkább, mint a neofunkcionalizáció) az ún. "duplikáció-degeneráció-komplementáció" jelensége, amire akkor van lehetőség, ha az eredeti gén (még megduplázódása előtt) több funkciót is ellátott. Ilyenkor a két új kópia feloszthatja egymás közt ezeket és ezzel együtt olyan mutációkat szedhetnek össze, amelyek a másik (komplementer) másolat által ellátott funkciókat befolyásolnák (s így az eredeti gén számára is károsak lennének), de a saját feladatok ellátására nincsenek hatással.
Új gének keletkezésének egyik jól bevált módja a génduplikáció, amikor a szóbanforgó DNS szakasza a genomnak valamilyen úton-módon, a sejtosztódás során megkétszereződik. Az így létrejövő extra másolat aztán viszonylag szabadon mutálódhat (hiszen az eredeti funkciót ellátja a másik kópia), ami leggyakrabban ahhoz vezet, hogy a gén működésképtelenné válik, de néha az is előfordul, hogy valamilyen új funkcióra tesz szert, ami ha előnyös az élőlény számára, jó eséllyel rögzülni fog (ezt nevezik a szakzsargonban "neofunkcionalizációnak").
Egy harmadik, a köztudatban kevésbé forgó lehetőség (bár egyáltalán nem biztos, hogy ritkább, mint a neofunkcionalizáció) az ún. "duplikáció-degeneráció-komplementáció" jelensége, amire akkor van lehetőség, ha az eredeti gén (még megduplázódása előtt) több funkciót is ellátott. Ilyenkor a két új kópia feloszthatja egymás közt ezeket és ezzel együtt olyan mutációkat szedhetnek össze, amelyek a másik (komplementer) másolat által ellátott funkciókat befolyásolnák (s így az eredeti gén számára is károsak lennének), de a saját feladatok ellátására nincsenek hatással.
Valami ilyesmi következett be a sörélesztő galaktóz lebontásért felelős egyik génjével, de mielőtt belevágnánk, hogy pontosan mi is történ(hetet)t, lássuk előbb, hogyan jön a képbe a galaktóz. A sörélesztő különböző cukrok, elsősorban szőlőcukor, vagyis glükóz lebontásával állítja elő a fennmaradásához szükséges energiát, miközben mellékesen alkoholt és szén-dioxidot állít elő (amit aztán mi, emberek ki is használunk, önös céljainkra ;-)). Ez a fermentáció folyamata, amihez azonban értelemszerűen szőlőcukorra van szükség.
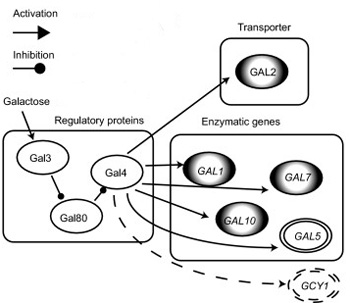 Néha azonban ínséges idők köszöntenek be, és a környéken nincs szőlőcukor, és az élesztőnek be kell érni más cukrokkal pl. galaktózzal. Ez utóbbinak a lebontásához más fajta géneket kell aktivizálni, de ugyanakkor fontos, hogy ezeket a géneket csak akkor kezdje átírni a sejt, ha valóban van galaktóz a közelben, különben csak fölösleges energiakidobás lenne a sok fehérje szintézise. Ennek megfelelően, a galaktóz lebontásért felelős enzimkaszkád tagjai egyaránt szigorú kontroll allatt vannak, s így nincs ez másképp az első lépésért felelős galaktokinázzal, azzal Gal1-el sem. A génátírást szabályozó régióban két fehérjéből, a GAL80-ból és GAL4-ből álló ún. transzkripciós represszor komplexek kapcsolódnak a DNS-hez (négy helyen is), alapesetben szigorúan letiltva a gének átírását.
Néha azonban ínséges idők köszöntenek be, és a környéken nincs szőlőcukor, és az élesztőnek be kell érni más cukrokkal pl. galaktózzal. Ez utóbbinak a lebontásához más fajta géneket kell aktivizálni, de ugyanakkor fontos, hogy ezeket a géneket csak akkor kezdje átírni a sejt, ha valóban van galaktóz a közelben, különben csak fölösleges energiakidobás lenne a sok fehérje szintézise. Ennek megfelelően, a galaktóz lebontásért felelős enzimkaszkád tagjai egyaránt szigorú kontroll allatt vannak, s így nincs ez másképp az első lépésért felelős galaktokinázzal, azzal Gal1-el sem. A génátírást szabályozó régióban két fehérjéből, a GAL80-ból és GAL4-ből álló ún. transzkripciós represszor komplexek kapcsolódnak a DNS-hez (négy helyen is), alapesetben szigorúan letiltva a gének átírását.
Ha a sejtbe galaktóz kerül, azt egy másik fehérje, a GAL3 érzékeli, és ilyenkor lefejti a GAL80 molekulákat a DNS-ről. Az egyedül ott maradó Gal4 pedig fordít egyet a köpönyegén, funkciót vált és elkezdi elősegíteni az enzimek átíródását (vázlatosan és GAL3 nélkül ugyanez itt is látható).
A nevezéktan egy kicsit zavaró, hiszen a folyamat összes tagja Galx néven fut, de ez esetben, pl. a Hox génektől eltérően, a közös előtag nem szerkezetbeli hasonlóságot jelöl, hanem azt, hogy ugyanannak a molekuláris útvonalnak a tagjairól beszélünk. Egyetlen kivétel van, ahol csak a Gal3 és Gal1 páros, amelyek ugyan teljesen más funkcióval rendelkeznek, mégis elég sok szerkezetbeli hasonlóságot mutatnak. Ez persze nem a véletlen műve, hanem egy korábbi genom duplikációjé, amely egy ősi génből kettőt "csinált".
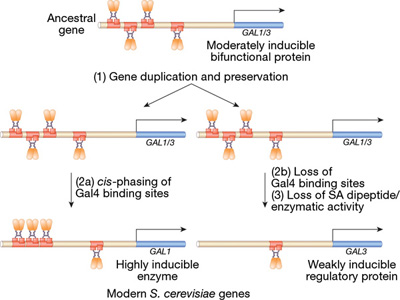 Ezt onnan is tudjuk, hogy egy szegről-végről rokon laktóz élesztőben (Kluyveromyces lactis), amelyik nem esett át genomduplikáción, a két funkciót egy és ugyanaz a gén látja el, a KlacGal1. A sörélesztőben a jelátvitelben jeleskedő GAL3 elvesztette ugyan az enzimatikus funkcióját, de a jelek szerint mindez inkább csak egy mellékszál, a valódi változások a génduplikáció után, a szabályozó régióban következtek be. Itt ugyanis a Gal3 már csak egyetlen GAL4 kötőhellyel büszkélkedhet, aminek következményeként, ha nincs galaktóz a rendszerben, akkor is átíródik egy kicsit, de ha lesz is galaktóz, akkor csak egy kicsit növekedik az átíródás üteme.
Ezt onnan is tudjuk, hogy egy szegről-végről rokon laktóz élesztőben (Kluyveromyces lactis), amelyik nem esett át genomduplikáción, a két funkciót egy és ugyanaz a gén látja el, a KlacGal1. A sörélesztőben a jelátvitelben jeleskedő GAL3 elvesztette ugyan az enzimatikus funkcióját, de a jelek szerint mindez inkább csak egy mellékszál, a valódi változások a génduplikáció után, a szabályozó régióban következtek be. Itt ugyanis a Gal3 már csak egyetlen GAL4 kötőhellyel büszkélkedhet, aminek következményeként, ha nincs galaktóz a rendszerben, akkor is átíródik egy kicsit, de ha lesz is galaktóz, akkor csak egy kicsit növekedik az átíródás üteme.
Mivel a GAL3 feladata, hogy érzékelje a galaktóz jelenlétét, értelemszerűen csak úgy működhet, ha alapból is van valamennyi fehérje a sejtben. Épp ezért az említett szabályozás igazából egy viszonylag optimális működést biztosít: mindig lesz elég fehérje a sejtben, hogy a jelátvitelt beindítsa. Ezzel szemben a Gal1 "mindent vagy semmit" elven működik, ha nincs galaktóz, alig észlelhető, de a legkevesebb galaktóz hatására közel ezerszeresére ugrik a koncentrációja.
A KlacGal1 promoterében, akárcsak a fent említett sörélesztő Gal1-nél, négy GAL4 kötő DNS szakasz van, de az első három (amelyek szorosan egymás mellett vannak), a sörélesztő Gal1 promoterétől eltérően nem teszi lehetővé, hogy a GAL4 fehérjék a spirál ugyanazon oldalán kössenek a DNS-hez. Mivel ez azt okozza, hogy a három GAL80-GAL4 komplex fizikailag kicsit távolabb csúszik egymástól, azzal is jár, hogy alap- (azaz galaktózmentes) állapotban a KlacGal1 transzkripció-gátlása nem annyira tökéletes mint a Gal1-é (bár még mindig jobb mint a Gal3-é). Ha felidézzük a Gal3 funkcióját (amit, ne feledjük, itt ugyancsak a KlacGal1 lát el), ez nem értelmetlen. De egyszersmind az is érezhető, hogy két funkció (a Gal3 ekvivalens jelátvivő, ill. a Gal1 ekvivalens enzimatikus) ellenkező irányba húzná a gén evolúcióját: a jelátvivő molekulából jobb ha kicsit több lenne, az enzimatikusból jobb, ha kicsit kevesebb, alapállapotban.
Magyarán a KlacGal1 mindkét feladata szempontjából szuboptimális állapotban van Kluyveromyces lactis-ban, de megváltozása bármelyik irányba, a másik funkcióját veszélyeztetné. Így aztán az optimalizálásra csak a génduplikáció után kerülhetett sor, amikor a két új gén feloszthatta egymás közt az eredeti feladatokat (lásd csatolt ábra).
Hittinger CT, Carroll SB (2007) Gene duplication and the adaptive evolution of a classic genetic switch. Nature 449: 677-681