Ha csak annyit írnék, hogy a posztban egy erősen szociális fajról lesz szó, melynek tagjai az egymással való érintkezéshez a kommunikáció egy igen fejlett formáját használják, és amely faj Afrikából (több hullámban) kiindulva hódította meg a világot, akkor valószínűleg sokan a Homo sapiens-re gondolnának, nem minden logikát nélkülözve, hiszen a felsorolt attribútumok egyike sem mondható el túl sok élőlényről, hát még a kombinációjuk.
Ha csak annyit írnék, hogy a posztban egy erősen szociális fajról lesz szó, melynek tagjai az egymással való érintkezéshez a kommunikáció egy igen fejlett formáját használják, és amely faj Afrikából (több hullámban) kiindulva hódította meg a világot, akkor valószínűleg sokan a Homo sapiens-re gondolnának, nem minden logikát nélkülözve, hiszen a felsorolt attribútumok egyike sem mondható el túl sok élőlényről, hát még a kombinációjuk.
De ezúttal mégsem a(z anatómiailag) modern emberről lesz szó, hanem egy sokkal kisebb és törékenyebb élőlényről, a háziméhről (aka. Apis mellifera) - ez azonban csak még elképesztőbbé teszi a fenti (rövid) leírást. És persze azt is hiba lenne elhallgatni, hogy a hasonlóságok azért inkább csak érintőlegesek és még ahol fel is fedezhetőek, ott is lényegesek a különbségek.
Ha csak annyit írnék, hogy a posztban egy erősen szociális fajról lesz szó, melynek tagjai az egymással való érintkezéshez a kommunikáció egy igen fejlett formáját használják, és amely faj Afrikából (több hullámban) kiindulva hódította meg a világot, akkor valószínűleg sokan a Homo sapiens-re gondolnának, nem minden logikát nélkülözve, hiszen a felsorolt attribútumok egyike sem mondható el túl sok élőlényről, hát még a kombinációjuk.
De ezúttal mégsem a(z anatómiailag) modern emberről lesz szó, hanem egy sokkal kisebb és törékenyebb élőlényről, a háziméhről (aka. Apis mellifera) - ez azonban csak még elképesztőbbé teszi a fenti (rövid) leírást. És persze azt is hiba lenne elhallgatni, hogy a hasonlóságok azért inkább csak érintőlegesek és még ahol fel is fedezhetőek, ott is lényegesek a különbségek.
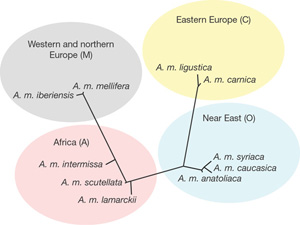 Vegyük például az eredet esetét. Ugyan az Afrikában őshonos háziméh két nagy hullámban vette be Eurázsiát (az első Ázsiát és K-Európát kolonizálta, a másik pedig Ny-Európát), de a hasonlóság itt meg is szűnik. (Megj.: az amerikai elterjedés szintén két hullámban történt; mindkettő részben emberi koordinációval. Előbb az "európai" alfajt vitték be 1622 táján a telepesek, ezt azonban a 20. században gyors ütemben kiszorította az agresszívebb afrikai alfaj, amelyet 1956-ban telepítettek be Brazíliába.) Hiszen a Homo genusszal ellentétben, melynek korai evolúciója is a "fekete kontinensre" lokalizálódik, a méhek ősei nem Afrikában szelték a levegőt. Ennek a jele az, hogy a ma ismert tíz Apis fajból kilenc Ázsiában őshonos, és a legrégebbi méh-fosszília is itt került elő. Ez utóbbi (Mellitosphex burmensis) egy burmai borostyánkő-bányában bukkant a felszínre és az Apoidea csoport egy mára már kihalt ágát képviseli; amolyan igazi "átmeneti fosszília", hiszen szépen keverednek benne a darázs- és méhvonások. Ráadásul azt igazolja, hogy a méhek kialakulása (~100 millió éve) az Alsó -Krétára tehető, vagyis igen közel a nyitvatermők kialakulásához - ami igen logikusnak tűnik, tekintve hogy utóbbiak megjelenése azt is jelentette, hogy egy remek új táplálékforrás tűnt fel a színtéren.
Vegyük például az eredet esetét. Ugyan az Afrikában őshonos háziméh két nagy hullámban vette be Eurázsiát (az első Ázsiát és K-Európát kolonizálta, a másik pedig Ny-Európát), de a hasonlóság itt meg is szűnik. (Megj.: az amerikai elterjedés szintén két hullámban történt; mindkettő részben emberi koordinációval. Előbb az "európai" alfajt vitték be 1622 táján a telepesek, ezt azonban a 20. században gyors ütemben kiszorította az agresszívebb afrikai alfaj, amelyet 1956-ban telepítettek be Brazíliába.) Hiszen a Homo genusszal ellentétben, melynek korai evolúciója is a "fekete kontinensre" lokalizálódik, a méhek ősei nem Afrikában szelték a levegőt. Ennek a jele az, hogy a ma ismert tíz Apis fajból kilenc Ázsiában őshonos, és a legrégebbi méh-fosszília is itt került elő. Ez utóbbi (Mellitosphex burmensis) egy burmai borostyánkő-bányában bukkant a felszínre és az Apoidea csoport egy mára már kihalt ágát képviseli; amolyan igazi "átmeneti fosszília", hiszen szépen keverednek benne a darázs- és méhvonások. Ráadásul azt igazolja, hogy a méhek kialakulása (~100 millió éve) az Alsó -Krétára tehető, vagyis igen közel a nyitvatermők kialakulásához - ami igen logikusnak tűnik, tekintve hogy utóbbiak megjelenése azt is jelentette, hogy egy remek új táplálékforrás tűnt fel a színtéren.
Az "eredethez" hasonlóan a nyelv esetében is felületes a hasonlóság. Nincsenek azonos nyelvtani szabályok, hasonló hangképzés, mondatszerkezet, azaz semmi, amit általában az emberi beszéddel szoktunk összefüggésbe hozni.
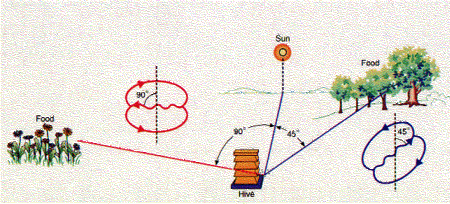 De ennek ellenére mai ismereteink szerint a főemlősökön kívül a méhek az egyetlen szimbolikus nyelvvel (lásd alább) rendelkező faj - ez pedig, tekintve, hogy agyuk mindössze egymillió idegsejtből áll (azaz kb. öt nagyságrenddel kevesebb, mint a mienk), nem rossz teljesítmény.
De ennek ellenére mai ismereteink szerint a főemlősökön kívül a méhek az egyetlen szimbolikus nyelvvel (lásd alább) rendelkező faj - ez pedig, tekintve, hogy agyuk mindössze egymillió idegsejtből áll (azaz kb. öt nagyságrenddel kevesebb, mint a mienk), nem rossz teljesítmény.
(A "szimbolikus nyelv" - természetesen - Karl von Frisch híres méhtáncát jelenti, amely segítségével a nektár/pollen gyűjtésből hazaérkező dolgozók társaikkal közlik az élelemforrás helyét.)
Méhekről a legtöbb embernek mindenekelőtt a kaptár (pontosabban a benne lakó raj) jut eszébe - na és persze a méz -, ez a hihetetlenül szofisztikált mikrotársadalom, amely lakóinak altruizmusa révén már-már egy szupraindividuális élőlényként működik. A rajt alkotó méhek (királynő, dolgozók, herék) egymás közeli rokonai, ami egyrészt elméleti síkon remek magyarázatot kínál az altruizmus jelenlétére (hiszen, még ha az egyes dolgozók nem is szaporodnak, így végül mégiscsak hozzájárulnak saját génjeik továbbéléséhez), másrészt azonban egy igencsak praktikus és valós problémát vet: a genetikailag túlontúl egyforma kolónia túlélési esélyei lényegesen rosszabbak a valós világ néha kiszámíthatatlan körülményei között (ennek megfelelően a természetben a diverzebb méhrajok sokkal gyorsabban tudnak növekedni, mint homogén társaik).
Egy genetikus szemszögéből azonban szinte kétségtelenül az a legérdekesebb, hogy egy viszonylag hasonló genetikai állomány hogyan kódolja a három igencsak különböző méh-életutat. A legegyszerűbb a herék, azaz hímek esete, akik társaikkal ellentétben haploidok, vagyis minden kromoszómából csak egyet hordoznak. Ez egyben oka is nemiségüknek, mert a haploid kromoszómakészlet azt jelenti, hogy a complementary sex determiner (csd) génből csak egy(fajta) verziót hordoznak, ez pedig a méh szex-determinációs rendszerben hím életutat eredményez - kvázi életük egyetlen célja a szaporodás.
A királynő és dolgozók esete bonyolultabb, hiszen mindannyian diploidok. Az egyetlen különbség, ami eldönti, hogy mi lesz a petéből, egyáltalán nem genetikai ok, hanem nagyon is környezeti: a táplálék minőségétől függ. Ez azonban nemcsak méretbeli és korbeli különbséget okoz (a királynő kétszer akkora, mint egy átlagos dolgozó és kétszer annyi ideig is él), de számos anatómiai különbséget is. A legevidensebb a petefészkek mérete - királynő: nagy; dolgozó: kisebb vagy még kisebb -, a hátsó lábak alakja - a dolgozóknál itt egy speciális "nektár kosár" alakul ki, de említhető a fullánk alakja is, ami a nőstényeknél nélkülözi a dolgozók kis kampóját (így csípésük is kevésbé kellemetlen).
A szemfülés kérdés persze az, hogy miért kapnak egyes peték több/jobb kaját mint a többiek. A helyzet az, hogy a királynő az "agykontroll" egészen különleges formáját alkalmazva igazgatja kis uradalmát. Egy különleges anyagot, ún. queen mandibular pheromone-t (QMP) termel, amelynek hatására a dolgozók más táplálékot adnak a petéknek, illetve szorgalmasabban végzik lakhelyük tisztítását. Épp ezért, ha elpusztul a királynő - vagyis nincs több QMP a levegőben-, alattvalói mintha "zombi álomból" ébrednének, hirtelen teljesen másképp kezdenek viselkedni. Egyrészt elkezdenek gourmet élelmet adni a még meglevő petéknek (így több királynő-jelölt is kikel, akik majd egymás között lejátsszák, hogy ki legyen a leendő uralkodó), ill. maguk is elkezdenek petéket rakni - de, mivel ők maguk sosem párosodtak, ezek haploid egyedek lesznek, azaz hímek. A tartós QMP dózis valószínűleg nem lehet kellemes és a királynőnek igyekeznie is kell, hogy ez ne tudatosuljon környezetében; erre látszik legalább is utalni a hormon leszedáló jellege: a feromon hatása alatt élő dolgozók nem képesek asszociatív tanulásra (pl. egy szagérzetet, a vele együtt adott enyhe áramütéshez kapcsolni).
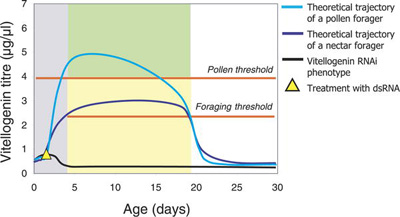 A méhek viselkedés-repertoárjának extra csavarja, hogy durva megközelítésben dolgozóból is három van: a lépet és kaptárt rendbentartó, valamint a pollent és nektárt gyűjtők. Előbbi és két utóbbi alapvetően ugyanannak az életútnak a különböző állomásait jelenti, hiszen kikelése után a méh kb. két hétig a kaptárban végzi a házimunkát, ezután pedig élte fennmaradó részét gyűjtögetéssel tölti. Azt, hogy éppen melyik "viselkedési program" fut, egy hormon (a juvenilis hormon - JH) és egy szikanyag fehérje, a vitellogenin koncentrációja határozza meg. A kettőnek ellentétes hatása van és egymást is kölcsönösen gátolják. A JH ösztönzi a gyűjtögetésre való áttérést, de igazából a vitellogenin koncentráció a mérvadó - csak, ha utóbbi egy küszöbérték alá csökken ("foraging threshold"), akkor következhet be a viselkedésváltás. A kezdeti hetek vitellogenin szintje azt is eldönti, hogy a dolgozó majd pollent vagy nektárt fog gyűjteni ("pollen forager" ill. "nectar forager"), ha kirepül: magas koncentráció esetén előbbi, relatíve alacsony esetén pedig utóbbi következik be. (Ezzel a mechanizmussal magyarázható, hogy ha a vitellogenin szintet mesterségesen lenullázzuk a fiatal méhben, akkor egyáltalán nem sertepertél a kaptárban, hanem azonnal kiszáll nektárt keresni.)
A méhek viselkedés-repertoárjának extra csavarja, hogy durva megközelítésben dolgozóból is három van: a lépet és kaptárt rendbentartó, valamint a pollent és nektárt gyűjtők. Előbbi és két utóbbi alapvetően ugyanannak az életútnak a különböző állomásait jelenti, hiszen kikelése után a méh kb. két hétig a kaptárban végzi a házimunkát, ezután pedig élte fennmaradó részét gyűjtögetéssel tölti. Azt, hogy éppen melyik "viselkedési program" fut, egy hormon (a juvenilis hormon - JH) és egy szikanyag fehérje, a vitellogenin koncentrációja határozza meg. A kettőnek ellentétes hatása van és egymást is kölcsönösen gátolják. A JH ösztönzi a gyűjtögetésre való áttérést, de igazából a vitellogenin koncentráció a mérvadó - csak, ha utóbbi egy küszöbérték alá csökken ("foraging threshold"), akkor következhet be a viselkedésváltás. A kezdeti hetek vitellogenin szintje azt is eldönti, hogy a dolgozó majd pollent vagy nektárt fog gyűjteni ("pollen forager" ill. "nectar forager"), ha kirepül: magas koncentráció esetén előbbi, relatíve alacsony esetén pedig utóbbi következik be. (Ezzel a mechanizmussal magyarázható, hogy ha a vitellogenin szintet mesterségesen lenullázzuk a fiatal méhben, akkor egyáltalán nem sertepertél a kaptárban, hanem azonnal kiszáll nektárt keresni.)
A HTB-gyűjtögető váltásnak következményeként a méhek génjeinek közel 40%-ában figyelhető meg expresszióváltozás. Hogy ezek közül melyek az igazán fontosak, azt még éppen csak kapisgáljuk, de remélhetőleg a tavaly elkészült háziméh-genom segítséget nyújt majd. (Ez utóbbi egyik plusz hozománya, hogy extra bizonyítékot nyújtott arra, amit már a hidra genom kapcsán feszegettem: a genetikusok legelső háziállata, a Drosophila genomja sok szempontból inkább kivételt képez, és aligha tekinthető átlagos "rovargenomnak". Például a kromoszómák végén levő ún. telomér szekvenciájuk igazából tök egyedi, még a méhekben hasonló ahhoz is, amit a gerincesekben találunk. Továbbá úgy tűnik, hogy míg a méhekben funkcionális DNS-metiláció figyelhető meg - bár ennek szerepe még nem tisztázott -, a muslicákban nem biztos, hogy van ilyen.)
A komplex társadalmi berendezkedés természetesen szinte tálcán kínálja a kérdést, hogy egy ilyen összetett viselkedés lérejöhetett-e evolúciósan. S bár nincs még tökélete bizonyítékunk, elég valószínűnek és logikusnak tűnő elméletünk már van. Ennek egyik fontos eleme, hogy a háziméhek minden viselkedésmintázata kezdetleges vagy kevésbé kezdetleges formában felfedezhető más, magányos darázs- és méhfajokban. Ezeknél általában a nőstény egyedül készít lakhelyet utódainak, amit azok fejlődése alatt rendben tart, majd pedig a peték kikelése után, azokat táplálja. Azaz igazából csak a feladatok hatékony felosztása jelent meg a háziméheknél (ill. velük párhuzamosan és tőlük függetlenül még más darázs, hangya és méh fajokban is): a királynő a feladat első részére "koncentrál", míg a dolgozók a többi részére. Ez persze azt jelenti, hogy az ősi viselkedésmintázat egyes elemei genetikailag szétválaszthatóak, erre pedig még nincs bizonyíték (részben azért mert a magányos méhfajok tanulmányozása nem az a tudományterület, amibe hagyományosan sok pénzt és energiát feccelnek).
De, tekintve a méhek gazdasági jelentőségét, ill. viszonylagos veszélyeztetettségét, nem tűnik kérdésesnek, hogy a méhek még hosszú ideig intenziv kutatómunka alanyai lesznek.
Vergoz V, Schreurs HA, Mercer AR (2007) Queen pheromone blocks aversive learning in young worker bees. Science 317: 384-386.
Mattilla HR, Seeley TD (2007) Genetic diversity in honey bee colonies enhances productivity and fitness. Science 317: 362 - 364.
Toth AL, Robinson GE (2007) Evo-devo and the evolution of social behavior. Trends Genet. 23: 334-341.
Page RE Jr, Amdam GV (2007) The making of a social insect: developmental architectures of social design. Bioessays 29: 334-343.
Nelson CM, Ihle KE, Fondrk MK, Page Jr. RE, Amdam GV (2007) The Gene vitellogenin Has Multiple Coordinating Effects on Social Organization. PLoS Biol 5(3): e62 doi:10.1371/journal.pbio.0050062
Honeybee Genome Sequencing Consortium (2006) Insights into social insects from the genome of the honeybee Apis mellifera. Nature 443: 931-949.
Whitfield CW, Behura SK, Berlocher SH, Clark AG, Johnston JS, Sheppard WS, Smith DR, Suarez AV, Weaver D, Tsutsui ND (2006) Thrice out of Africa: ancient and recent expansions of the honey bee, Apis mellifera. Science 314: 642-645.
Wang Y, Jorda M, Jones PL, Maleszka R, Ling X, Robertson HM, Mizzen CA, Peinado MA, Robinson GE (2006) Functional CpG methylation system in a social insect. Science 314: 645-647.
Poinar GO Jr, Danforth BN (2006) A fossil bee from Early Cretaceous Burmese amber. Science 314: 614.