


 Ha a nemek mögött levő genetikai faktorokról beszélünk, elsősorban az X és Y kromoszómára gondolunk. Pedig az emlősökre olyan jellemző "XY" szexdeterminációs rendszer (egyébként vannak kivételek, azokról majd máskor) csak logikájában tekinthető viszonylag elterjedtnek az állatvilágban, mechanizmusában egyáltalán nem.
Ha a nemek mögött levő genetikai faktorokról beszélünk, elsősorban az X és Y kromoszómára gondolunk. Pedig az emlősökre olyan jellemző "XY" szexdeterminációs rendszer (egyébként vannak kivételek, azokról majd máskor) csak logikájában tekinthető viszonylag elterjedtnek az állatvilágban, mechanizmusában egyáltalán nem.
Gondolom, fölösleges hangsúlyozni, de az emlősök esetében a homogametikus, azaz két db. X kromoszómával rendelkező egyedek nőstények lesznek, a heterogametikus, azaz X és Y kromoszómát egyaránt hordozó társaik pedig hímek. Azonban más magzatburkos gerincesekben (az egyszerűség kedvéért maradjunk most ilyen szűk körben) ez nem feltétlenül van így: pl. madarakban éppen fordított a helyzet. Itt a homogamétás nem tagjai (ezeket ZZ-vel jelöljük) válnak hímekké, míg a heterogamétás (ZW) társaik nőstényekké. És ez még viszonylag kis különbségnek számít, mert hüllők és kétéltűek között egyaránt lelünk olyan fajokat, amelyekben az XY-, vagy a ZW-rendszer alapján működik a szexdetermináció, vagy éppen környezeti hatások alapján (a klasszikus példa itt a krokodilokra és tengeri teknősökre jellemző hőmérsékletfüggés - a kicsit magasabb hőmérsékleten fiúk kelnek ki a tojásokból, míg alacsonyabban lányok). Sőt, ó irgalom atyja ne hagyj el, kétéltűeknél van, amikor egyetlen faj (!!) különböző populációi különböző szexdeterminációs technikát alkalmaznak, vagy éppen valami környezeti hatásra megváltoztatják a nemüket.
Ha a nemek mögött levő genetikai faktorokról beszélünk, elsősorban az X és Y kromoszómára gondolunk. Pedig az emlősökre olyan jellemző "XY" szexdeterminációs rendszer (egyébként vannak kivételek, azokról majd máskor) csak logikájában tekinthető viszonylag elterjedtnek az állatvilágban, mechanizmusában egyáltalán nem.
Gondolom, fölösleges hangsúlyozni, de az emlősök esetében a homogametikus, azaz két db. X kromoszómával rendelkező egyedek nőstények lesznek, a heterogametikus, azaz X és Y kromoszómát egyaránt hordozó társaik pedig hímek. Azonban más magzatburkos gerincesekben (az egyszerűség kedvéért maradjunk most ilyen szűk körben) ez nem feltétlenül van így: pl. madarakban éppen fordított a helyzet. Itt a homogamétás nem tagjai (ezeket ZZ-vel jelöljük) válnak hímekké, míg a heterogamétás (ZW) társaik nőstényekké. És ez még viszonylag kis különbségnek számít, mert hüllők és kétéltűek között egyaránt lelünk olyan fajokat, amelyekben az XY-, vagy a ZW-rendszer alapján működik a szexdetermináció, vagy éppen környezeti hatások alapján (a klasszikus példa itt a krokodilokra és tengeri teknősökre jellemző hőmérsékletfüggés - a kicsit magasabb hőmérsékleten fiúk kelnek ki a tojásokból, míg alacsonyabban lányok). Sőt, ó irgalom atyja ne hagyj el, kétéltűeknél van, amikor egyetlen faj (!!) különböző populációi különböző szexdeterminációs technikát alkalmaznak, vagy éppen valami környezeti hatásra megváltoztatják a nemüket.
Szóval bonyolult ez. Látható, hogy a genetikai szexdetermináció többször, több-módon kialakult, ami nem teljesen véletlen. Egy állandó környezetű világban szép és jó a hőmérsékletfüggő megoldást alkalmazni: ilyenkor a homokpadban megbúvó tojásrakás közepéből kisfiúk, széléből meg kislányok kelnek ki. Azonban elég egy hirtelen jött klímaingadozás és máris gebasz van: néhány fok ide, vagy oda és csak fiúk/lányok látnak napvilágot. Ez pedig nem éppen adaptív stratégia. Azaz a genetikai determináció (legyen az XY vagy ZW alapú) vitathatatlanul előnyös, hiszen az ideális fele-fele arányt lehet vele belőni.
A legelfogadottab feltételezések szerint éppen ezzel magyarázható, hogy a magzatburkosok előszeretettel cserélték le a (feltehetően ősi) környezeti faktorokon alapuló megoldást. Azért beszélhetünk a látszólag különböző rendszerek közös eredetéről, mert a végső szereplők minden fajban ugyanazok: Dmrt1, Sox9, WT1, SF1, AMH - csupa olyan fehérje, amely az emberi nem-meghatározásban is kulcsszerepet játszik. Úgy tűnik, hogy csak az egész folyamatot szabályozó kulcsgén mibenléte különbözik fajról fajra. (Ennek az lehet az oka, hogy az ősi, többlépéses környezetihatás-függő rendszert különböző fajok különböző módon szakították meg, beiktatva egy genetikai főszabályozót.) Annyiban azonban közös az XY és ZW logikája, hogy ez a kulcsgén azon a nemi kromoszómán helyezkedik el, amelyik a heterozigótás nem kialakulásához szükséges (Y ill. W).
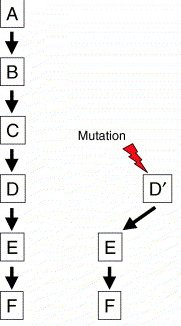 Az egész folyamat kezdetén a két nemi kromoszómapár egyáltalán nem különbözik egymástól, valójában egy teljesen hétköznapi autoszóma-párt alkotnak, amelyeken a nem-meghatározásért felelős gének (vagy azokat befolyásoló faktorok) is helyet foglalnak (a jobb oldali ábrán, amely a Schartl cikk egyik illusztrációja nyomán készült, ezt D jelöli). Egy adott ponton azonban a kromoszómapár egyik tagján az egyik ilyen gén összeszed egy domináns mutációt, aminek következtében akkor is aktív lesz, ha egyébként erre nem kap utasítást. Így annak az utódnak a neme, amelyik ezt a kromoszómát örökli, függetlenné válik a környezettől (az emlősök esetében ez a hím, ill. Y kromoszóma). Ekkor a rendszer még elég labilis, hiszen egy "rosszul sikerült" rekombináció meiotikus sejtosztódás során visszaállíthatja az eredeti állapotot. Azonban ha mielőtt ez bekövetkezne, a domináns gén régiójában egy inverzió, vagy más kromoszómaszintű átalakulás zajlik le, többé nem áll fenn a rekombináció veszélye, hiszen megszűnnek a homológ régiók. Az allél megmenekült és folytatódhat az új mechanizmus tuningolása. Ennek keretében olyan "apróságok" kell kialakuljanak, mint annak a biztosítása, hogy a homogamétás egyedek ellenkező neműekké váljanak, mint a (domináns allélt hordozva determinációjukat már bebiztosított) heterogamétásak.
Az egész folyamat kezdetén a két nemi kromoszómapár egyáltalán nem különbözik egymástól, valójában egy teljesen hétköznapi autoszóma-párt alkotnak, amelyeken a nem-meghatározásért felelős gének (vagy azokat befolyásoló faktorok) is helyet foglalnak (a jobb oldali ábrán, amely a Schartl cikk egyik illusztrációja nyomán készült, ezt D jelöli). Egy adott ponton azonban a kromoszómapár egyik tagján az egyik ilyen gén összeszed egy domináns mutációt, aminek következtében akkor is aktív lesz, ha egyébként erre nem kap utasítást. Így annak az utódnak a neme, amelyik ezt a kromoszómát örökli, függetlenné válik a környezettől (az emlősök esetében ez a hím, ill. Y kromoszóma). Ekkor a rendszer még elég labilis, hiszen egy "rosszul sikerült" rekombináció meiotikus sejtosztódás során visszaállíthatja az eredeti állapotot. Azonban ha mielőtt ez bekövetkezne, a domináns gén régiójában egy inverzió, vagy más kromoszómaszintű átalakulás zajlik le, többé nem áll fenn a rekombináció veszélye, hiszen megszűnnek a homológ régiók. Az allél megmenekült és folytatódhat az új mechanizmus tuningolása. Ennek keretében olyan "apróságok" kell kialakuljanak, mint annak a biztosítása, hogy a homogamétás egyedek ellenkező neműekké váljanak, mint a (domináns allélt hordozva determinációjukat már bebiztosított) heterogamétásak.
S bár a kromoszómaátrendeződés sikeresen kimentette kulcsgénünket a rekombináció karmaiból, ennek ára is van: rekombináció hiányában felhalmozódnak a mutációk, így ez a kromoszóma az idők folyamán fokozatosan elsatnyul, (a kulcsgén kivételével) egyre több gén válik működésképtelenné. Ezért olyan satnya és génszegény saját Y kromoszómánk is.
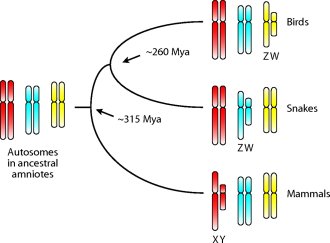 Na, akkor lassan végre elértünk oda, hogy miért is volt szükség erre a röpke bevezetőre. Mivel a kígyók esetében is a ZW determináció működik, és a kígyók és madarak viszonylag közeli rokonok, első látásra elképzelhetőnek tűnik, hogy valami távoli közös ősben alakult ki egy közös szexdeterminációs rendszer. Ennek természetesen az lenne a feltétele, hogy a kígyó és madár Z (ill. W) kromoszómák valódi homológok legyenek, vagyis ugyanazon gének helyezkedjenek el rajtuk. A helyzet azonban nem ez, a kígyók Z kromoszómáján levő gének a madarak 2-es autoszómáján találhatók és vice versa, a csirkék Z kromoszómájára jellemző markerek a kígyó autoszómákon vannak. (Hasonlóan, az emberi X kromoszóma sem a madár nemi kromoszómákkal rokon, hanem az 1. és 4. autoszómákkal.) Vagyis a madarak és kígyók nagyon hasonló mechanizmusú szexdeterminációs mechanizmusa egymástól függetlenül alakult ki az évmilliók során.
Na, akkor lassan végre elértünk oda, hogy miért is volt szükség erre a röpke bevezetőre. Mivel a kígyók esetében is a ZW determináció működik, és a kígyók és madarak viszonylag közeli rokonok, első látásra elképzelhetőnek tűnik, hogy valami távoli közös ősben alakult ki egy közös szexdeterminációs rendszer. Ennek természetesen az lenne a feltétele, hogy a kígyó és madár Z (ill. W) kromoszómák valódi homológok legyenek, vagyis ugyanazon gének helyezkedjenek el rajtuk. A helyzet azonban nem ez, a kígyók Z kromoszómáján levő gének a madarak 2-es autoszómáján találhatók és vice versa, a csirkék Z kromoszómájára jellemző markerek a kígyó autoszómákon vannak. (Hasonlóan, az emberi X kromoszóma sem a madár nemi kromoszómákkal rokon, hanem az 1. és 4. autoszómákkal.) Vagyis a madarak és kígyók nagyon hasonló mechanizmusú szexdeterminációs mechanizmusa egymástól függetlenül alakult ki az évmilliók során.
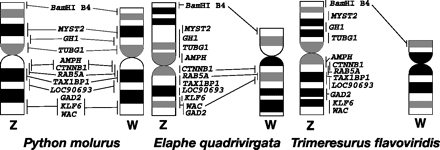
A kígyók történetében hab a tortán, hogy a különböző fajokban nyomon követhetjük a heterogamétás W kromoszóma elsatnyulását (ez nem unikum, a madaraknál is lehet látni, de azért szép példa). Az ősi pitonfélék családjába tartozó burmai piton (Python molurus) esetében a W kromoszómán ugyanazon a géneket fedezhetjük fel, mint a Z-n. A lényegesebben fiatalabb, viperafélék csoportjába tartozó habu (Trimeresurus flavoviridis) esetében, a W nemcsak hogy szemmel láthatólag kisebb, de el is vesztette a vizsgált 11 gén mindegyikét. A köztes négycsíkú patkánysikló (Elaphe quadrivirgata) W kromoszómája egy átmeneti állapotot képvisel méretben is és genetikai információ szempontjából is: a 11 génből 3 még mindig megtalálható benne.
Már csak annyi a kérdés, hogy mi lehet a szexdetermináció kulcsgénje a kígyók esetében, mert ez egyelőre homályban maradt.
(Az első kép a www.indigosnakes.com oldalról származik.)
Matsubara, K, Tarui, H, Toriba, M, Yamada, K, Nishida-Umehara, C, Agata, K, Matsuda, Y (2006) Evidence for different origin of sex chromosomes in snakes, birds, and mammals and step-wise differentiation of snake sex chromosomes. PNAS 103: 18190-18195.
Vallender, EJ, Lahn, BT (2006) Multiple independent origins of sex chromosomes in amniotes. PNAS 103: 18031-18032.
Schartl, M (2004) Sex chromosome evolution in non-mammalian vertebrates. Curr Opin Gen Dev 14: 634-641.




