 Nem igazán múlhat el hét valamilyen Hox gén sztori nélkül (max. az én hibámból ;-)), szóval lássuk, mi is keltett az elmúlt napokban nagy hírverést biológus berkekben (s mint a végén látni fogjuk, nemcsak ott).
Nem igazán múlhat el hét valamilyen Hox gén sztori nélkül (max. az én hibámból ;-)), szóval lássuk, mi is keltett az elmúlt napokban nagy hírverést biológus berkekben (s mint a végén látni fogjuk, nemcsak ott).
A post témájaként szolgáló kísérlet szellemes, ugyanakkor technikailag egyáltalán nem egyszerű, s bár számos fontos dolgot sikeresen példáz, a nemzetközi és hazai média kicsit túllőtt a célon a cikk ismertetésekor.
Zanzásítva, a kísérlet során azt vizsgálták, hogy mennyire fedhetnek át a különböző Hox-klaszterekben elhelyezkedő paralóg gének funkciói, illetve mi az, ami miatt mégis különbözik a szerepük a szervezet fejlődése során. Mivel ez a nem szakmabeliek számára elég meredekül hangozhat, lássuk csak egy kicsit bővebben, miről is van szó.
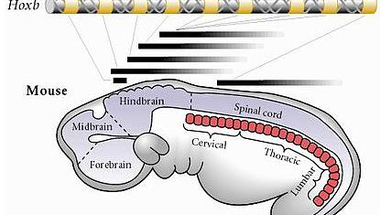
A blog visszatérő motívumaként szuperáló Hox gének a test hosszanti tagolásáért felelnek és ebben igen fontos szerepe van a kollineáris kifejeződésüknek, ami röviden annyit tesz (mint a mellékelt ábra is mutatja), hogy a gének kromoszómán megfigyelhető sorrendje egyben azt is tükrözi, hogy az embrió fejlődése során miként fejeződnek ki az embrionális hossztengely mentén. Ma már egyre jobb képünk van arról, miként is alakulhatott ki ez a bűvös, kollineáris Hox-klaszter (a közeljövőben még lesz erről szó), de azért még rengeteg megválaszolatlan kérdés is van. Pl. hogy miért is küzdenek látszólag az állkapcsos gerincesek a bőség zavarával, hiszen különböző genomduplikációk okán és nyomán négy darab Hox klaszter van genetikai állományukban, azaz kis túlzással olyan, mintha minden egyes Hox génből négy kópia lenne (valójában nem mindegyikből van négy, de min. kettő azért előfordul). Holott az evolúció szereti kiszórni a redundáns funkciójú géneket, vagyis az a fogas kérdés merül fel, hogy a látszólag egyforma Hox gének valójában teljesen egyformák-e.
Hogy lassan a lényegre térjek, az ominózus cikk szerzői a Hox1 paralógokat vizsgálják. Pontosabban a utóagyban kifejeződő (lásd fenti ábra) és annak fejlődésében szerepet játszó Hoxa1 és Hoxb1 funkcióit hasonlították össze (azért csak ezt a kettőt, mert Hoxc1 nincs, ez a gén elveszett a harmadik Hox-klaszterből, a Hoxd1 pedig nem játszik szerepet az emlős-utóagy fejlődésében). A két génről annyi volt tudott, hogy mutációik az utóagy szegmenseinek, a rhomboméráknak helytelen fejlődését okozzák, amiből aztán mindenféle - sokszor letális - galiba adódik (a Hoxb1 mutációja egyes arcizmok mozgatóidegeinek elsorvadását okozza). Illetve arra is rájöttek korábban, hogy a Hoxa1 részt vesz a Hoxb1 átíródásának szabályozásában.
A kísérlet során "egyszerűen" kicserélték a két gént, a Hoxa1 fehérje kódoló szekvenciájának helyére a Hoxb1 megfelelő szekvenciáját szúrták, illetve vice versa (a létrejövő mutáns, a klasszikus nomenklatúra értelmében a Hoxa1B1(g)/B1(g)Hoxb1A1(g)/A1(g) nevet nyerte a keresztségben). A keletkező kisegerek vígan élték világukat, azaz a felületes szemlélő könnyen arra a a következtetésre juthatott, hogy a két fehérje mindössze 49%-os hasonlóságuk ellenére (ami azért a kulcsfontosságú DNS kötő régióban 85%-os) teljesen azonos funkciót lát el.
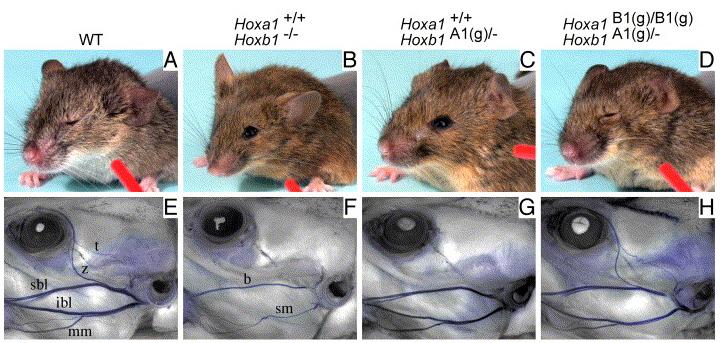
De mialatt a két gén különböző allélkombinációit vizsgálgatták, néhány dolog azért szemet szúrt a kísérletezőknek. Ugyanis amíg a Hoxa1+/+Hoxb1+/- genotípusú egerek jóformán megkülönböztethetetlenek nem mutáns (azaz Hoxa1+/+Hoxb1+/+) társaiktól, addig a Hoxa1+/+Hoxb1A1(g)/- egérfiókák a Hoxb1 mutánsokhoz (Hoxa1+/+Hoxb1-/-) hasonló arcbénulással küzdenek (az ilyen egerek, ha egy szívószálon keresztül az arcukba fújunk, nem hunyorítanak), ugyancsak a mozgató arcideg hiánya miatt (lásd a fenti ábra második sorát). Érdekes módon a Hoxa1B1(g)/B1(g)Hoxb1A1(g)/- genotípusú egerek többsége viszont normális e téren, azaz a kísérletek erősen azt sugallták, hogy a két gén azért mégsem teljesen egyenrangú.
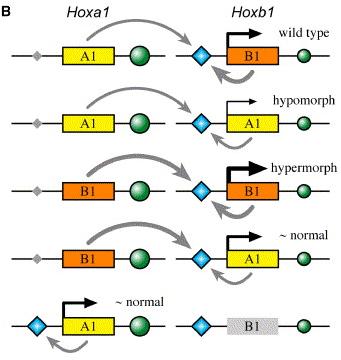 Hogy megértsük, miben rejlik a különbség, ahhoz néhány szót kell ejtsek a Hox1 gének szabályozásáról. Alapvetően két szabályozó elem fontos, az egyik, amely a retinolsav gradiens mentén aktiválja a Hoxa1 expresszióját (RARE - retinoic acid regulatory element) - ezt a jobb oldali ábrán egy zöld kör jelöli -, a másik pedig egy önszabályozó elem (ARE - auto regulatory element), amely segítségével a Hoxa1 aktiválja a Hoxb1-et illetve utóbbi önmaga átíródását is elősegíti - ez pedig a kék rombusz. A Hoxa1 után egy igen aktív RARE van, de az előtte levő ARE elsatnyult, a Hoxb1 esetében pedig éppen fordított a helyzet. Summa summarum, a kísérletek során arra derült fény, hogy a Hoxb1 sokkal jobban képes az ARE elemen keresztüli aktiválásra, mint a Hoxa1. Ezért néznek ki viszonylag normálisan a Hoxa1B1(g)/B1(g)Hoxb1A1(g)/A1(g)egerek, ahol végső soron megfelelő mennyiségű Hox1 géntermék jön létre a felcserélt gének kölcsönhatása révén, illetve ezért arcbénulásosak a Hoxa1+/+Hoxb1A1(g)/- társaik - itt a Hoxb1 helyén levő Hoxa1 gén hemizigóta állpotban nem képes saját átírását eléggé felpörgetni.
Hogy megértsük, miben rejlik a különbség, ahhoz néhány szót kell ejtsek a Hox1 gének szabályozásáról. Alapvetően két szabályozó elem fontos, az egyik, amely a retinolsav gradiens mentén aktiválja a Hoxa1 expresszióját (RARE - retinoic acid regulatory element) - ezt a jobb oldali ábrán egy zöld kör jelöli -, a másik pedig egy önszabályozó elem (ARE - auto regulatory element), amely segítségével a Hoxa1 aktiválja a Hoxb1-et illetve utóbbi önmaga átíródását is elősegíti - ez pedig a kék rombusz. A Hoxa1 után egy igen aktív RARE van, de az előtte levő ARE elsatnyult, a Hoxb1 esetében pedig éppen fordított a helyzet. Summa summarum, a kísérletek során arra derült fény, hogy a Hoxb1 sokkal jobban képes az ARE elemen keresztüli aktiválásra, mint a Hoxa1. Ezért néznek ki viszonylag normálisan a Hoxa1B1(g)/B1(g)Hoxb1A1(g)/A1(g)egerek, ahol végső soron megfelelő mennyiségű Hox1 géntermék jön létre a felcserélt gének kölcsönhatása révén, illetve ezért arcbénulásosak a Hoxa1+/+Hoxb1A1(g)/- társaik - itt a Hoxb1 helyén levő Hoxa1 gén hemizigóta állpotban nem képes saját átírását eléggé felpörgetni.
Na, akkor ezek után jöjjön az a kísérlet, amire rápörgött a világsajtó. Legvégül arra lettek kíváncsiak a kutatók, hogy mi lesz, ha a Hoxb1 helyett egyszerűen az előtte levő aktív ARE-t szúrják be a Hoxa1 elé (s közben a Hoxb1-et szépen elrontják). Gondolom, már nem meglepő, hogy a Hoxa1ARE(g)/ARE(g)Hoxb1-/-egerek teljesen normálisnak tűntek.
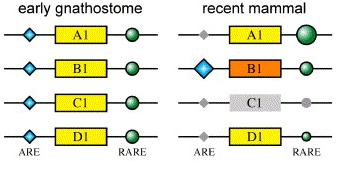 Na, akkor hogy is jön ez az egész az evolúcióhoz. A különböző genomszekvenálgatások és más örökítőanyag-vizsgálatok egyik hozadéka, hogy ma eléggé úgy tűnik, hogy az állkapcsos gerincesek közös ősében mindenféle kromoszómaduplikációk során egyszerre négy, egyenrangú Hox1 gén lehetett jelen, amelyek éppen megfelelő mennyiségű Hox1 fehérjét hoztak létre retinolsav- és önreguláció során, hogy az utóagy és a különböző utóagyi idegek normálisan kialakuljanak . Az emlősökhöz vezető evolúciós úton aztán az egyik Hox1 gén (a Hoxc1) elveszett, egy másik (Hoxd1) szabályozása elsatnyult, a maradék kettő pedig ellenkező irányban variálgatta RARE és ARE szabályozó elemeinek érzékenységét. Ennek eredményeképpen összességében a Hox1 fehérjeszint alig változott, bár kombinatorikusan teljesen más úton jön létre.
Na, akkor hogy is jön ez az egész az evolúcióhoz. A különböző genomszekvenálgatások és más örökítőanyag-vizsgálatok egyik hozadéka, hogy ma eléggé úgy tűnik, hogy az állkapcsos gerincesek közös ősében mindenféle kromoszómaduplikációk során egyszerre négy, egyenrangú Hox1 gén lehetett jelen, amelyek éppen megfelelő mennyiségű Hox1 fehérjét hoztak létre retinolsav- és önreguláció során, hogy az utóagy és a különböző utóagyi idegek normálisan kialakuljanak . Az emlősökhöz vezető evolúciós úton aztán az egyik Hox1 gén (a Hoxc1) elveszett, egy másik (Hoxd1) szabályozása elsatnyult, a maradék kettő pedig ellenkező irányban variálgatta RARE és ARE szabályozó elemeinek érzékenységét. Ennek eredményeképpen összességében a Hox1 fehérjeszint alig változott, bár kombinatorikusan teljesen más úton jön létre.
Azaz a médiában használt "visszafordították egérben az evolúciót" cím abban az értelemben teljesen félrevezető, hogy sohasem létezett olyan egér, amelynek egyetlen Hox1 génje lett volna, ellenben a kísérletsorozat nagyon szépen példázza azt, hogy genomduplikációk után az egyes gének a szabályozó elemeik változtatgatásával feloszthatnak maguk között kezdetben közös feladatokat, azaz szubfunkcionalizálódhatnak.
Tvrdik P, Capecchi M. (2006) Reversal of Hox1 gene subfunctionalization in the mouse. Dev Cell 11(2): 239-250.