Szinte napra pontosan Darwin közelmúltbeli szülinapjára tette fel honlapjára a Nature azt a cikket, amit az egykori természettudós minden bizonnyal nagy élvezettel olvasott volna, hiszen egyik kedvenc állatcsoportjáról, a galapagosi pintyekről van benne szó.
Szinte napra pontosan Darwin közelmúltbeli szülinapjára tette fel honlapjára a Nature azt a cikket, amit az egykori természettudós minden bizonnyal nagy élvezettel olvasott volna, hiszen egyik kedvenc állatcsoportjáról, a galapagosi pintyekről van benne szó.
Pontosabban a cikk tárgya 120 (igen, százhúsz), a legkülönbözőbb fajokhoz tartozó egyed genomszekvenciájának meghatározása és azok összevetése.
Ez gigászi mennyiségű adat és bőven lehet benne érdekes dolgokra lelni, mind arra vonatkozóan, hogy miként zajlott ezeknek a fajoknak a speciációja, mind arról, hogy az ikonikus csőrformák változása milyen DNS-szintű változásokra vezethető vissza.
Az egyik érdekes eredmény az a klasszikus pinty-törzsfa részleges átrajzolásához vezetett, mégpedig azért mert a hegyescsőrű földipinty (Geospiza difficilis) különböző populációiról kiderült, hogy azok hasonló kinézete nem annyira a közvetlen rokonságnak, hanem sokkal inkább a konvergens evolúciónak köszönhetők - magyarán nem ugyanazon fajhoz tartoznak. Ezen kívül gyakori a hibridizáció is az egyes fajok között - ami, bár elsőre ellentmondani látszik a szigorúan értelmezett biológiai fajfogalomnak, amely szerint külön fajok nem hozhatnak létre szaporodóképes utódokat, valójában csak azt igazolja, hogy a fajok kialakulása során van egy "szürke zóna", ahol a két elkülönülő populáció már sok jellegben különbözik, de még időről időre van köztük kapcsolat.
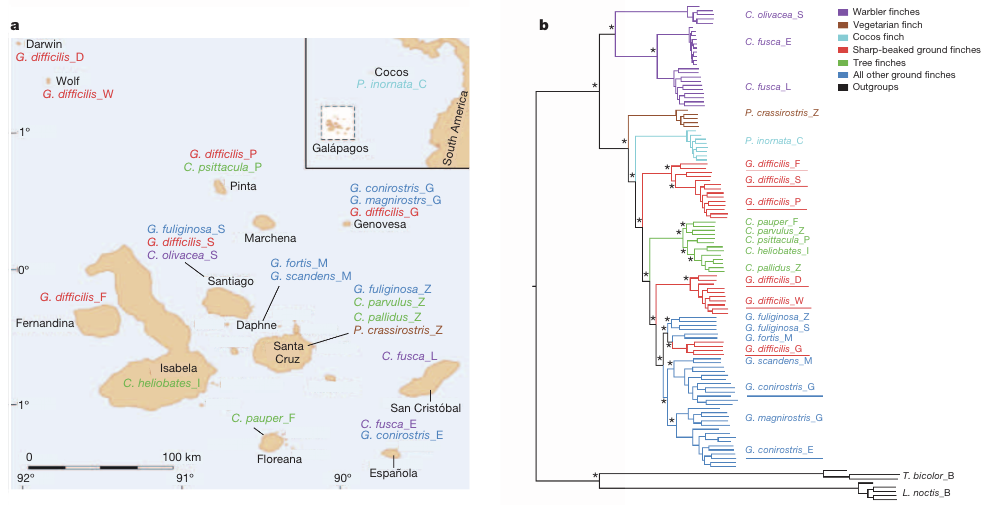 A tompa és hegyes csőrű fajok genetikai diverzitásának összevetése arra nyújtott apropót, hogy olyan tipikus különbségeket keressenek, amelyek pintyfajok "cégérének" kialakulását és relatíve gyors evolúciós változását magyarázhatják. Korábbi vizsgálatok már fellebbentették a fátylat arról, hogy egyrészt a calmodulin (CALM) nevű kálcium-szenzor fehérje, valamint a BMP-jelátviteli útvonal is felelőssek ennek a fenotípusnak az alakulásáért, de valószínűsíthető volt, hogy ez még nem a teljes kép.
A tompa és hegyes csőrű fajok genetikai diverzitásának összevetése arra nyújtott apropót, hogy olyan tipikus különbségeket keressenek, amelyek pintyfajok "cégérének" kialakulását és relatíve gyors evolúciós változását magyarázhatják. Korábbi vizsgálatok már fellebbentették a fátylat arról, hogy egyrészt a calmodulin (CALM) nevű kálcium-szenzor fehérje, valamint a BMP-jelátviteli útvonal is felelőssek ennek a fenotípusnak az alakulásáért, de valószínűsíthető volt, hogy ez még nem a teljes kép.
A teljes genom összvetések egyrészt visszaigazolták, hogy a CALM-t kódoló gén különböző alléljai lehetnek jelen a tompa illetve hegyes csőrű pintyekben, ugyanakkor a korábban hírbe-hozott BMP4 gén esetében nem leltek olyan, szelekciós nyomás alatt levő genetikai eltérést, ami relevánsnak tünnének, így valószínűbb, hogy a BMP4 expresszióját befolyásoló transzkripciós faktorok között vannak allélikus különbségek a különböző csőrű madarak között.
Apropó transzkripciós faktorok: a genom-összevetések egyik fontos eredménye, hogy kiderült, a két csőrtípus közti különbségek jelentős része az ALX1 génhez kapcsolható. Az ALX1 egy homeobox transzkripciós faktor, amely a gerinces arc fejlődésében fontos szerepet játszik és amelynek a humán mutációi arc-deformációhoz vezetnek.
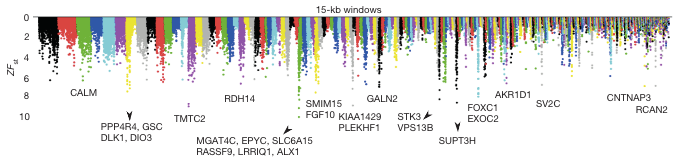
A tanulmány során sikerült egy tompa (blunt - B) és egy hegyes (pointed - P) csőrért felelős ún. ALX1 haplotípust azonosítani, aminek segítségével nagyon nagy bizonyossággal lehet jósolni, pusztán DNS alapján, hogy annak a gazdája milyen csőrrel rendelkezett.
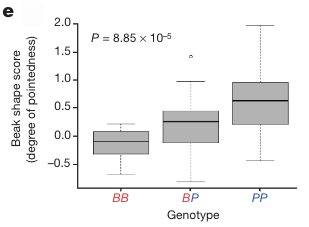 Sőt, a homozigóta BB, illetve PP, valamint a heterozigóta BP haplotípusok nem csak fajok között rendelkeztek nagy prediktív erővel, hanem egy konkrét fajon belül is tükrözték, hogy a hordozójuk milyen csőrrel rendelkezik.
Sőt, a homozigóta BB, illetve PP, valamint a heterozigóta BP haplotípusok nem csak fajok között rendelkeztek nagy prediktív erővel, hanem egy konkrét fajon belül is tükrözték, hogy a hordozójuk milyen csőrrel rendelkezik.
A szóbanforgó faj ez esetben a Geospiza fortis, amely esetében a galapagosi pintyek két leghíresebb megfigyelője, Peter és Rosemary Grant pár évvel ezelőtt leírta, hogy viszonylag gyors éghajlatváltozásokhoz is gyorsan tudtak alkalmazkodni.
Ezt, a mostani adatok szerint, az tette lehetővé, hogy a G. fortis pont azon kevés fajok egyike, ahol a tompa és hegyes csőrért felelős ALX1 B és a P haplotípus egyszerre jelen van a populációkban - vagyis nagyszerű genetikai "alapanyaga" volt a természetes szelekciónak.
(via Nature News)
Lamichhaney S, Berglund J, Almén MS, Maqbool K, Grabherr M, et al. (2015) Evolution of Darwin/'s finches and their beaks revealed by genome sequencing. Nature http://dx.doi.org/10.1038/nature14181