Úgy tűnik, hogy a galapagosi pintyek nehezen megunható kutatási "tárgyat" jelentenek. Alig fél hónappal a Grant házaspár Science cikke után, amelyben egy fontos evolúciós elméletet, a jelleg kiszorítást igazolták ezeken a kis madarakon, itt van egy újabb szösszenet (ezúttal a Nature-ben jelent meg), amelyben Clifford Tabin csoportja, újból a nyomába ered annak, hogy milyen molekuláris okokra ezethető vissza a csőrtípusok közötti eltérés.
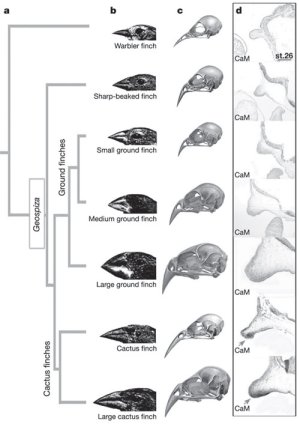 Mondom újból, hiszen Tabin nevéhez fűződik az a cikk is, amiről egyszer már tettem említést: ott a bmp4 nevű gén expressziójának erőssége és az egyes pintyfajok csőrének robusztussága (átmérőjének nagysága) között mutattak ki összefüggést. Azonban a bmp4 (és egyáltalán, a különböző BMP-k) kipróbálása szinte triviális volt, ismerve jól dokumentált általános hatását a csontozat fejlődésére. Ennek a megközelítésnek természetesen az a nagy hátránya, hogy annak alapján próbál egy folyamat szempontjából fontos géneket találni, hogy mit tudunk azok hatásáról egyébként. Azaz nem igazán alkalmas olyan faktorok nyakoncsípésére, amelyeket korábban még nem hoztak kapcsolatba a csontfejlődéssel. Ráadásul ez a klasszikus módszer egyáltalán nem adott választ arra, hogy mitől hegyesebb egyes galapagosi pinty (Geospiza) fajok csőre, mint más fajoké.
Mondom újból, hiszen Tabin nevéhez fűződik az a cikk is, amiről egyszer már tettem említést: ott a bmp4 nevű gén expressziójának erőssége és az egyes pintyfajok csőrének robusztussága (átmérőjének nagysága) között mutattak ki összefüggést. Azonban a bmp4 (és egyáltalán, a különböző BMP-k) kipróbálása szinte triviális volt, ismerve jól dokumentált általános hatását a csontozat fejlődésére. Ennek a megközelítésnek természetesen az a nagy hátránya, hogy annak alapján próbál egy folyamat szempontjából fontos géneket találni, hogy mit tudunk azok hatásáról egyébként. Azaz nem igazán alkalmas olyan faktorok nyakoncsípésére, amelyeket korábban még nem hoztak kapcsolatba a csontfejlődéssel. Ráadásul ez a klasszikus módszer egyáltalán nem adott választ arra, hogy mitől hegyesebb egyes galapagosi pinty (Geospiza) fajok csőre, mint más fajoké.
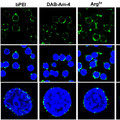
Hogy leszámoljanak a fenti a problémával, és egyúttal választ is kapjanak a csőrhegyesség misztikus kérdésére, Tabinék ún. DNS-chip-ek segítségével összehasonlították hegyes csőrű kaktuszpintyek (G. conirostris és G. scandens) és tompa csőrű földi pintyek (G. fortis és G. magnirostris) fejlődő embrióinak csőrkezdeményeiben kifejeződő géneket. A legkiemelkedőbb, közel ötszörös kifejeződési különbséget a calmodulin (CaM) nevű kálcium-szenzor fehérje génje mutatta. Mint azt a további vizsgálatok kimutatták, a CaM valóban szinte csak a katuszpintyek csőrében fejeződik ki kimutathatóan.
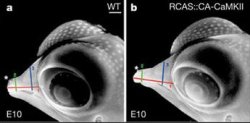 Persze ez a tény önmagában még nem biztosít ja a két jelenség közötti ok-okozati összefüggést, erre való azonban a laboratóriumi egerek szárnyas megfelelője, a csirkeembrió. S valóban, ha a CaM-hoz kapcsolódó jelátviteli útvonalt mesterségesem aktiválták (egy CaMKII molekula állanódan aktív formája - CA-CaMKII - segítségével), a csirkeembrióknak jól láthatóan és mérhetően hegyesebb lett a csőre, mint nem kezelt társaiknak.
Persze ez a tény önmagában még nem biztosít ja a két jelenség közötti ok-okozati összefüggést, erre való azonban a laboratóriumi egerek szárnyas megfelelője, a csirkeembrió. S valóban, ha a CaM-hoz kapcsolódó jelátviteli útvonalt mesterségesem aktiválták (egy CaMKII molekula állanódan aktív formája - CA-CaMKII - segítségével), a csirkeembrióknak jól láthatóan és mérhetően hegyesebb lett a csőre, mint nem kezelt társaiknak.
Vagyis jelen tudásunk segítségével már lehet egy elméletet felállítani arról, hogy alapvetően melyik két gén (bmp4 és CaM) járult hozzá nagyban a csőrforma gazdagság kialakulásához acsőr vastagságt illetve hegyességét szabályozva.
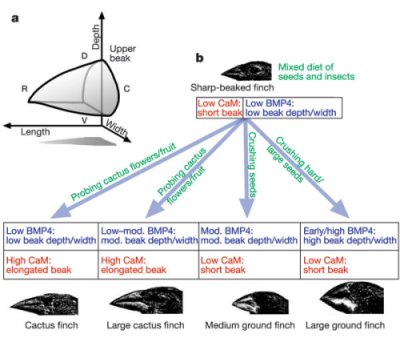
Az persze továbbra is nyitott kérdés, hogy a CaM expresszió változása általános-e a madár csőr hegyesség kialakulásában, vagy csak egy több alternatív útvonal közül. Ehhez azonban más fajok vizsgálatára is sort kell keríteni, a szerzők sugallnak is néhány ilyen "jelöltet", ezek a kolibrik, néhány parti madár nemzettség és a hawaii mézmadarak, amelyek maguk is igen változatos (de gyakran hegyes) csőrök tulajdonosai.
Abzhanov, A, Kuo, WA, Hartmann, C, Grant, BR, Grant, PR, Tabin, CJ (2006) The calmodulin pathway and evolution of elongated beak morphology in Darwin's finches. Nature 442: 563-567.