 Szélesebb rendszertani csoportunk közös jellemzője, hogy minden idetartozó faj a fejlődés valamelyik stádiumában egy jellegzetes képletet növeszt, a gerinchúrt.
Szélesebb rendszertani csoportunk közös jellemzője, hogy minden idetartozó faj a fejlődés valamelyik stádiumában egy jellegzetes képletet növeszt, a gerinchúrt.
Ez a rugalmas pálcára emlézetető, hatalmas vakuólumoktól feszülő sejtekből álló szerv ugyanúgy megtalálható egy emberi embrióban, mint egy hallárvában, vagy akár egy fiatal zsákállatban, és minden egyes egyedben az embriogenezis során egyfajta belső feszítő-vázként szolgál a szervezet számára. A fejlődés későbbi stádiumaiban gyakran fölösleges lesz, s ilyenkor vagy teljesen, nyomtalanul eltűnik (lásd zsákállatok), vagy beépül a létrejövő szövetekbe (így bennünk a porckorong belsejében lelhető fel a nyoma).
A gerinchúr olyan univerzális ismertető jegy is, amelynek köszönhetően biztosak lehetünk benne, hogy gerinchúrosok már 530 millió évvel ezelőtt is léteztek, mert ismerjük egy képviselőjüket a Pikaia-t.
Persze mindez aligha ad választ a kérdésre, hogy honnan ered ez a különös szerv? Mikor alakult ki, s főképpen vajon miből?
Nos, ha nem is minden fontos kérdésre válaszol Detlev Arendték új cikke, de néhány érdekes dologra fényt vet.
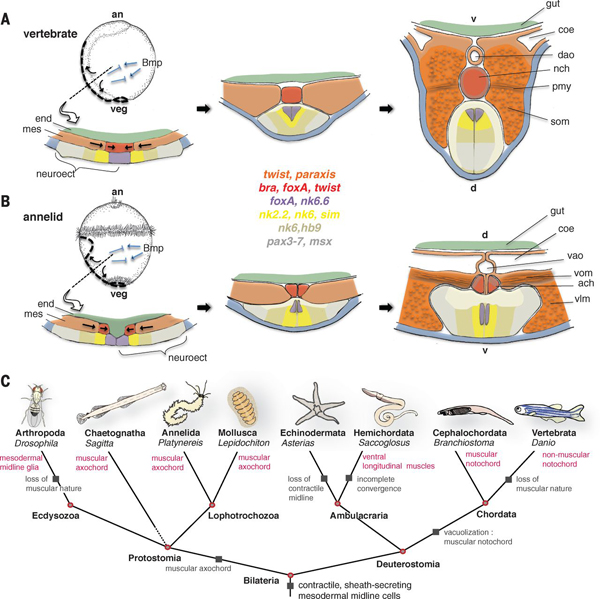
Detlev az elmúlt évek során hihetetlenül eredményesen feltérképezte a gerinces és gerinctelen csoportok fejlődése során megfigyelhető génexpressziós hasonlóságokat és egyebek mellett az ő munkájának köszönhetően tudjuk, hogy a két csoport közös ősének (a mitikus Urbilateriának) az idegrendszerét, vagy hát-hasi tengelyét ugyanazok a gének határozhatták meg, mint ma pl. egy halembrió idegrendszerét és hát-hasi tengelyét. A munkájában egy különleges tengeri gyűrűsféreg, a Platynereis dumerilii volt segítségére, ami az Urbilateria fejlődési titkainak feltárásán túl annak genomjáról is elárult néhány titkot: a Platynereis genom volt az első ahol rádöbbentünk, hogy az ősi többsejtű állatok genomjai igencsak gazdagok voltak intronokban és az az intronszegénység, amit ma néhány modellállatunkban (pl. C. elegans, vagy muslica) tapasztalunk, az ezekben a vonalakban specifikusan kialakult tulajdonság.
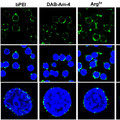
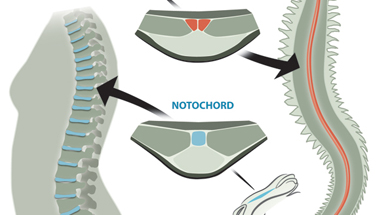
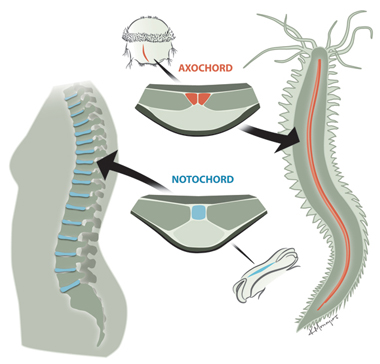
Arendték most is a Platynereis-ből indultak ki és azt keresték, hogy létezik-e olyan szerv, aminek ha funkciója és struktúrája nem is, de kialakulása és helyzete analógnak tekinthető a gerinchúréval. Végül leltek is egy ilyent, a test hasi oldalának közepén végigfutó, ún. mediális, ventrális, hosszanti izmot (mvlm). Ez ugyanúgy az idegrendszer alatt helyezkedik el, mint a gerinchúr (vagy szakzsargonban notochord) - itt az "alatt" azt jelenti, hogy a test mélyebb részei fele van -, és a kialakításában részt vevő sejtek a fejlődés során test két oldaláról érkeznek egy konvergens mozgás segítségével.
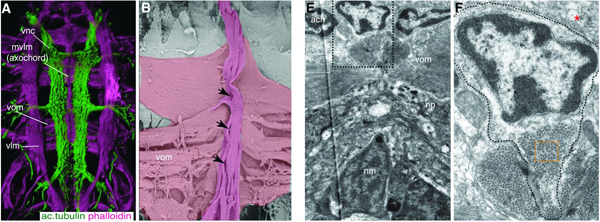
A kifejlett állatban az mvlm a ventrális ferde izomkötegbe (ventral oblique muscle - vom) feszül bele, és elektronmikroszkópos képe egyértelműen mutatja, hogy egy különleges izomsejtről van szó, amiben semmilyen fajta vakuólum nincs.
Mégis a puszta helyzeti és kialakulási analógia mellett Arendték azt is kimutatták, hogy azok a fejlődési génszabályozó hálózatok, amelyek nélkülözhetetlenek a notochord kialakításában, bekapcsolódnak az mvlm kialakulása során is. Ennek megfelelően a foxD, brachyury (bra), foxA, slit, hedgehog (hh) és noggin gének, amelyeket egytől egyig kifejeződnek a fejlődő notochordban, a kialakuló mvlm-ben is expresszálódnak.
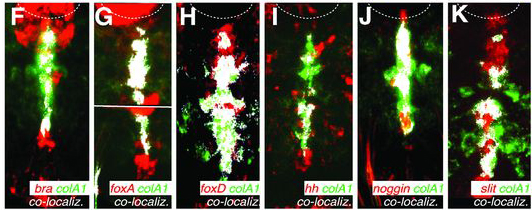
Ennek fényében Arendték, valószínűleg pontosan, arra a következtetésre jutottak, hogy a notochord és az mvlm a közös ős, az Urbilateria azonos szervéből alakulhatott ki, s ennek megfelelően az mvlm-t át is keresztelték "axochord"-nak.
Mivel a notochord és axochord sejtszintű felépítése nagyon-nagyon különbözik, nehéz megsaccolni, hogy mi is lehetett az ősi homológ struktúrának a kinézete és szerepe. Lehet, hogy egy egyszerű kötőszöveti sejtpopuláció volt, aminek egyetlen jellegzetessége, hogy a sejtjei valamilyen tokot képeztek maguk körül. De persze az sem zárható ki, hogy már akkor is valamilyen rugalmas szekrezetet alkottak ezek a sejtek és ez alakult az egyik fejlődési vonalban axochorddá, a másikban pedig notochorddá.
Bármi legyen is ez igazság, az azonban most már bizonyosnak látszik, hogy ez a különleges jelleg nem a semmiből jelent meg a mi csoportunkban, hanem nagyon is jól körülhatárolható sejtpopuláció hozta végül létre.
Lauri A, Brunet T, Handberg-Thorsager M, Fischer AH, Simakov O, et al. (2014) Development of the annelid axochord: insights into notochord evolution. Science 345: 1365-8.