Amit ma a különböző modellorganizmusok fejlődésbiológiájának genetikájáról tudunk, azt elsősorban az elmúlt bő 25 év genetikai screenjeinek köszönhetjük. Ha pedig ezekről beszélünk, akkor nem lehet nem megemlíteni Christiane Nüsslein-Volhard úttörő munkásságát, aki előbb Eric Wieschaus-al karöltve lenyomta a ma már tankönyvek lapjait tarkító, legendás ecetmuslica fejlődés-screent (amiért később, teljesen megérdemelten, megosztott Nobel-díjat kaptak), azóta pedig szinte folyamatosan, újabb és újabb zebrahal screenek felett bábáskodik. Legutóbbi nekifutásban, egy egyébként egyre telítődőbb mezőnyben, sikerült ismét eredetit alkotnia: kivételesen nem olyan mutációknak eredt a nyomába, amelyek a korai embrionális fejlődési rendellenességeket okoznak, hanem olyan mutációkat keres, amelyek a felnőtt jellegek változását okozzák. Halakról lévén szó, ez elsősorban pikkelyekkel és mintázatokkal kapcsolatos dolgokat jelent, ami unalmasan, az embertől végtelenül elvonatkoztatottnak hangzik talán, de persze nem az: a legtöbb pikkely-fejlődésben érintett génnek fontos szerepe van az emberi kültakaró származékok (pl. haj, fogak, izzadság mirigyek) kialakulásában is.
Amit ma a különböző modellorganizmusok fejlődésbiológiájának genetikájáról tudunk, azt elsősorban az elmúlt bő 25 év genetikai screenjeinek köszönhetjük. Ha pedig ezekről beszélünk, akkor nem lehet nem megemlíteni Christiane Nüsslein-Volhard úttörő munkásságát, aki előbb Eric Wieschaus-al karöltve lenyomta a ma már tankönyvek lapjait tarkító, legendás ecetmuslica fejlődés-screent (amiért később, teljesen megérdemelten, megosztott Nobel-díjat kaptak), azóta pedig szinte folyamatosan, újabb és újabb zebrahal screenek felett bábáskodik. Legutóbbi nekifutásban, egy egyébként egyre telítődőbb mezőnyben, sikerült ismét eredetit alkotnia: kivételesen nem olyan mutációknak eredt a nyomába, amelyek a korai embrionális fejlődési rendellenességeket okoznak, hanem olyan mutációkat keres, amelyek a felnőtt jellegek változását okozzák. Halakról lévén szó, ez elsősorban pikkelyekkel és mintázatokkal kapcsolatos dolgokat jelent, ami unalmasan, az embertől végtelenül elvonatkoztatottnak hangzik talán, de persze nem az: a legtöbb pikkely-fejlődésben érintett génnek fontos szerepe van az emberi kültakaró származékok (pl. haj, fogak, izzadság mirigyek) kialakulásában is.
A screen elején lelt pikkely-mutánsokról például kiderült, hogy ugyanazon, ectodyplasin függő útvonalban hibádznak, mint a pikkely nélküli tüskés pikók, vagyis kvázi laboratóriumi körülmények között, sikerült a természetes szelekciót reprodukálni.
 A csoport legutóbbi cikke, egy másik pikkely-mutánst vesz górcső alá, az ún. "tükör dániót" (spiegeldanio - spd), amely onnan kapta a nevét, hogy a halak kinézete nagyon emlékeztet a tükörpontyok (angolul mirror carp) pikkelytelenségére: amint a baloldali képen jól megfigyelhető, a pikkelyzetet vörösre festő ún. alizarin-festés mindkét esetben az oldalsó pikkelyek szinte teljes hiányát igazolta.
A csoport legutóbbi cikke, egy másik pikkely-mutánst vesz górcső alá, az ún. "tükör dániót" (spiegeldanio - spd), amely onnan kapta a nevét, hogy a halak kinézete nagyon emlékeztet a tükörpontyok (angolul mirror carp) pikkelytelenségére: amint a baloldali képen jól megfigyelhető, a pikkelyzetet vörösre festő ún. alizarin-festés mindkét esetben az oldalsó pikkelyek szinte teljes hiányát igazolta.
A mutáció térképezése aztán kiderítette, hogy egy növekedési faktor-receptort kódoló génben az fgfr1-ben következett be a mutáció: a fehérje aktivitásához fontos aminosav cserélődött ki, ezzel a receptor elvesztette az aktivitását.
Mivel mint szinte minden jelátviteli útvonal komponens, az Fgf1r is általában nélkülözhetetlen a fejlődéshez (mutáns egerek, de akár medaka halak is elpusztulnak az embriogenezis során), kisebb fejvakarást okozott, hogy a zebrahalakban miért csak ennyire enyhe hatást lehet megfigyelni. A teljes genom közelebbi vizsgálata, azonban kézenfekvő magyarázatot adott mindenre: egy génduplikáció következtében a zebrahalaknak két fgfr1 génje van van (fgfr1a és fgfr1b) és mivel a mutáció csak az előbbiben van, az utóbbi képes kompenzálni a hiányát. (Ha mindkét gén kiütjük, akkor az embrionális fejlődés során elpusztulnak a halak.)
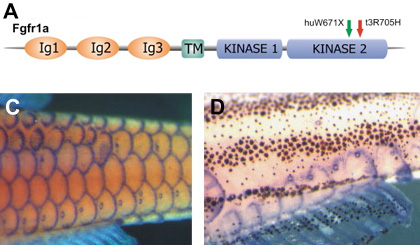 Persze ezek alapján jó kérdés, hogy az fgfr1b miért nem kompenzál a pikkelyekben is. A válasz azonban nem túl bonyolult: a két gén szinte mindig egy helyen fejeződik ki, de csak szinte - pont a pikkelyek jelentik a kivételt. Ezekben az fgfr1a jellegzetesen ott fejeződik ki, ahol a pikkely növekedése történne (lásd mellékelt ábrán mind a vadtípus (C), mind a spd (D) esetében), ellenben az fgfr1b nem található meg. Jól mutatja mindez, hogy a gén és genomduplikációk miként adhatnak táptalajt a szelekciónak: olyan gének esetében, amelyek számos jellegért felelnek, a duplikátum (vagy az eredeti) szabadon változhat, mert sokkal kevesebb baj származik belőle, s időnként akár valami jó is kisülhet a dologból.
Persze ezek alapján jó kérdés, hogy az fgfr1b miért nem kompenzál a pikkelyekben is. A válasz azonban nem túl bonyolult: a két gén szinte mindig egy helyen fejeződik ki, de csak szinte - pont a pikkelyek jelentik a kivételt. Ezekben az fgfr1a jellegzetesen ott fejeződik ki, ahol a pikkely növekedése történne (lásd mellékelt ábrán mind a vadtípus (C), mind a spd (D) esetében), ellenben az fgfr1b nem található meg. Jól mutatja mindez, hogy a gén és genomduplikációk miként adhatnak táptalajt a szelekciónak: olyan gének esetében, amelyek számos jellegért felelnek, a duplikátum (vagy az eredeti) szabadon változhat, mert sokkal kevesebb baj származik belőle, s időnként akár valami jó is kisülhet a dologból.
A "tükör" fenotípus molekuláris okának ismeretében, logikusnak tűnhet azon is elfilózni, hogy a tükörpontyokkal való hasonlóságnak, hasonló genetikai okai vannak-e.
Rövidre fogva a történetet: igen. A zebrahalakkal egyébként viszonylag közeli rokon tükörpontyok esetében az emberi szelekció, a jelek szerint kétszer, egymástól függetlenül, ugyanezt az fgfr1a gént "lelte meg". (Megj.: itt egy picit egyszerűsítek, mert a pontyokban egy extra genomduplikáció miatt két fgr1a gén van, fgfr1a1 és fgfr1a2, de eleve csak az előbbi működőképes, és ebben vannak a szóbanforgó mutációk is a tükörpontyok esetében.) Egyik esetben egy deléció miatt 37 aminosav hiányzik a fehérjetermékből (ami így persze működésképtelen lesz), a másik esetben pedig egy másik konzervatív aminosav megváltozása nullázza le a receptor funkcióját.
(A cikk magyar vonatkozása, hogy a tükörpontyok Keszthelyről, a Pannon Egyetem Georgikon karán tanító Bercsényi Miklóstól származnak.)
Rohner N, Bercsényi M, Orbán L, Kolanczyk ME, Linke D, Brand M, Nüsslein-Volhard C, Harris MP. (2009) Duplication of fgfr1 Permits Fgf Signaling to Serve as a Target for Selection during Domestication. Curr. Biol. doi:10.1016/j.cub.2009.07.065