

 Az evolúciót legjobban szemléltető fajokról alkotott szubjektív toplistámom, a galapagosi pintyek, a tüskés pikó és a nyírfaaraszoló lepkék mellett igen előkellő helyet foglal el egy mexikói lazacfaj is, az Astyanax mexicanus. (Kicsit offtopic kitérő: a nyírfaaraszolóról lehet elvétve olyan pletykákat hallani kreacionista berkekből, hogy valójában nem is szemléltetik jól az evolúciót, de nem kell felülni nekik: mint annyi másban, ebben is tévednek.)
Az evolúciót legjobban szemléltető fajokról alkotott szubjektív toplistámom, a galapagosi pintyek, a tüskés pikó és a nyírfaaraszoló lepkék mellett igen előkellő helyet foglal el egy mexikói lazacfaj is, az Astyanax mexicanus. (Kicsit offtopic kitérő: a nyírfaaraszolóról lehet elvétve olyan pletykákat hallani kreacionista berkekből, hogy valójában nem is szemléltetik jól az evolúciót, de nem kell felülni nekik: mint annyi másban, ebben is tévednek.)
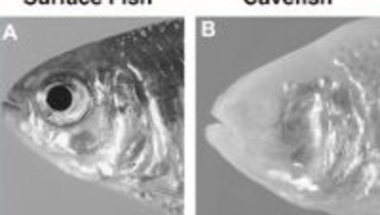
Az Astyanax-nak alapvetően két populációja ismert: egyik a felszíni tavakban él és teljesen hétköznap kinézetű (ez "surface fish" néven szerepel majd az ábrákon), a másik pedig a Yucatán félszigetet alkotó mészkőben képződött barlangok tavaiban lakik, teste pigmentszegény és első pillantásra úgy tűnik, hogy teljesen vak, hiszen egyáltalán nincs is szeme ("cavefish").
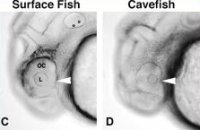 Az első látás azonban picit (de nem nagyon) csalóka. Ugyanis bár ezek a halak funkcionálisan kétségtelenül vakok, a szemük azért kialakul a fejlődés során. Egy egészen fiatal, egynapos embrióban (lásd baloldalt) még egészen jól látható a szem, bár már látszik, hogy nincs minden rendben vele: kicsit kisebb, és az alsó (ventrális) része szinte teljesen hiányzik. Később azonban fokozatosan elsatnyul, s bár nem tűnik el teljesen, a fejlődő lárva más szövetei körbenövik.
Az első látás azonban picit (de nem nagyon) csalóka. Ugyanis bár ezek a halak funkcionálisan kétségtelenül vakok, a szemük azért kialakul a fejlődés során. Egy egészen fiatal, egynapos embrióban (lásd baloldalt) még egészen jól látható a szem, bár már látszik, hogy nincs minden rendben vele: kicsit kisebb, és az alsó (ventrális) része szinte teljesen hiányzik. Később azonban fokozatosan elsatnyul, s bár nem tűnik el teljesen, a fejlődő lárva más szövetei körbenövik.
A kérdés tehát adott: mind molekuláris, mind evolúciós-adaptációs szemszögből, miért nincs szeme a barlangban élő Astyanax-oknak? Bár a teljes választ még nem ismerjük, elég sok mindent tudunk arról, hogy sejtsük hol kell keresgélni. Ráadásul a kérdés megválaszolását kétségtelenül segíti mind az a tény, hogy számos más halfaj (pl. a zebrahal és a medaka) már bejáratott fejlődésbiológiai kísérleti "nyúl" (vagyis általában elég sokat tudunk a halak fejlődéséről), mind az a fontos tény - amit nem is hangsúlyoztam az elején -, hogy a felszíni és barlangi lazacpopulációk vidáman keresztezhetőek egymással (ez megkönnyíti a mutáció(k) beazonosítását).
![]() Már az első kísérletek is azt sugallták, hogy a szem egészen korai embrionális fejlődésével nem stimmel valami. Amint arról minap írtam, a szemfejlődés egy fontos lépése, amikor a szemhólyag és a lencse-placod közti molekuláris összjáték eredményeképpen előbbiből kialakul a szemserleg, az utóbbi pedig szemlencseként lefűződik.
Már az első kísérletek is azt sugallták, hogy a szem egészen korai embrionális fejlődésével nem stimmel valami. Amint arról minap írtam, a szemfejlődés egy fontos lépése, amikor a szemhólyag és a lencse-placod közti molekuláris összjáték eredményeképpen előbbiből kialakul a szemserleg, az utóbbi pedig szemlencseként lefűződik.
A barlangi Astyanax-ok szemlencséje azonban pusztuló sejtekből áll, éppen ezért nem tud megfelelő viszontjelzéseket adni a formálódó szemserlegnek, ezért marad az viszonylag satnya. De ha, egy felszíni halembrió lencse-placodját átültetjük rgy vele egykorú barlangi társába, akkor utóbbiban, az átültetés helyén ép és egészésges, működő szem alakul ki ("transplant side")! (A "control side" azt mutatja, hogy mi lesz, ha egy másik barlangi Astyanax lencséjét ültetik át - semmi.)
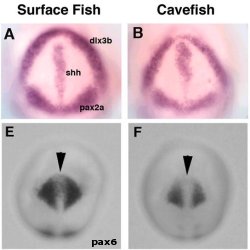 Hát ez egyszerű, gondolhatnánk, akkor tehát a szemlencse sejtjeivel van a baj. Na nem egészen. Az előbb is emlegetett molekuláris összjáték azt jelenti, hogy a lencse félrefejlődéséért, akár a szemhólyag is felelős lehet (ha nem küld megfelelő jeleket, a lencse sem fejlődik normálisan). És valóban, a jelek azt mutatják, hogy itt van a kutya elásva. Ugyanis a kétfele osztódott szemmező (lásd ismét a gerinces szemfejlődés postot) két, pax6 által jelölt oldala kisebb a barlangi lazacokban, mint a felszíniekben (E,F). A különbség nem tűnik drámainak, de, mint azt mindjárt látni fogjuk, ahhoz éppen elég, hogy a szóbanforgó változás oka legyen.
Hát ez egyszerű, gondolhatnánk, akkor tehát a szemlencse sejtjeivel van a baj. Na nem egészen. Az előbb is emlegetett molekuláris összjáték azt jelenti, hogy a lencse félrefejlődéséért, akár a szemhólyag is felelős lehet (ha nem küld megfelelő jeleket, a lencse sem fejlődik normálisan). És valóban, a jelek azt mutatják, hogy itt van a kutya elásva. Ugyanis a kétfele osztódott szemmező (lásd ismét a gerinces szemfejlődés postot) két, pax6 által jelölt oldala kisebb a barlangi lazacokban, mint a felszíniekben (E,F). A különbség nem tűnik drámainak, de, mint azt mindjárt látni fogjuk, ahhoz éppen elég, hogy a szóbanforgó változás oka legyen.
A szemmező változása a középvonal, pontosabban az ebben levő shh expressziós terület kiszélesedése miatt következik be (A, B). Ismétlem, a különbség mindössze néhány sejtben mérhető: amíg a felszíni halakban kb. 6 sejtsor vastagságú a középvonal Shh+ területe, addig a barlangi populációkban 10 sejtsorról van szó.
 Lehetséges-e, hogy ez a minimális különbség okozza az elég drasztikus különbséget a felnőtt halak között? Ennek eldöntésére egy kis extra Shh-t jutattak felszíni Astyanax-ok embriójának egyik oldalába. Az eredmény magért beszél: már az egynapos embrióknak is lényegesen kisebb a szeme a "kezelés" oldalán (H), a felnőtt halakban, pedig nagyon hasonlóan a barlangi rokonokhoz, a bőr benövi a szemet, azaz a szóbanforgó oldalon a halak kvázi vakok lesznek (J).
Lehetséges-e, hogy ez a minimális különbség okozza az elég drasztikus különbséget a felnőtt halak között? Ennek eldöntésére egy kis extra Shh-t jutattak felszíni Astyanax-ok embriójának egyik oldalába. Az eredmény magért beszél: már az egynapos embrióknak is lényegesen kisebb a szeme a "kezelés" oldalán (H), a felnőtt halakban, pedig nagyon hasonlóan a barlangi rokonokhoz, a bőr benövi a szemet, azaz a szóbanforgó oldalon a halak kvázi vakok lesznek (J).
Bár a mutációt még konkrétan nem sikerült kiszúrni, a legtöbben valamilyen szabályozó régió megváltozására fogadnak. (A legtöbb fejlődésben fontos gén nagyon sok helyen hat az embriogenezis során, éppen ezért valószínű, hogy egy ilyen kicsi és lokalizált változás nem egy gén teljes működésképtelenségére vezethető vissza.) A legérdekesebb az egészben az, hogy az eddigi genetikai vizsgálatok azt mutatják, hogy a bár ez az Astyanax faj legalább öt külön vonalon vált vakká (magyarán minimum öt olyan barlangi populáció ismert, amelyek különböző mutációk miatt jöttek létre), a shh kifejeződés erősödése, ill. a lencse elpusztulása minden esetben bekövetkezett.
És ezzel elértünk a szakmai berkekben sokáig legvitatottabb kérdéshez: mi az evolúciós oka a szem hiányának? Egyrészt elképzelhető, hogy a szemfejlődésben bekövetkezett malőr annak tudható be, hogy a hal amúgy sem használta a látószervét, így a szem kialakulásában fontos gének (szabályozó szekvenciái) szabadon elromolhattak, hiszen természetes szelekció nem szűrte ki a megvakult példányokat, és azok később valahogy elterjedtek (ez a "neutrális mutáció" elmélete). Másrészt az is egy lehetséges magyarázat, hogy azok a változások, amelyek a kvázi vakságot okozzák, egyben olyan más változásokhoz is vezetnek, amelyek kifejezetten előnyösek a hal számára (ez pedig az "adaptív evolúció" elmélete).
Jelen ismereteink alapján utóbbinak áll a zászló. Ugyanis nyilvánvaló, hogy a Shh expressziója nem elromlott, hanem éppen hogy túlságosan hatékony lett. Másrészt azok a gének, amelyek csak a szemfejlődésben szerepelnek (ilyenből is akad néhány), s így a neutrális mutációk legkézenfekvőbb célpontjai lehetnének, továbbra is működőképesek a vaklazacban és át is íródnak a fejbe besüllyedt, elsorvadt, más szövetek által körbenőtt szemben. A valószínűbb magyarázat az (és erre számos bizonyíték van), hogy a Shh expresszió megnövekedése egyben erősíti a szaglótevékenységet és egy robosztusabb álkapocsszerkezet kialakulását. Ezek pedig olyan tulajdonságok, amelyek nyilvánvalóan előnyösek lehetnek a barlangok sötétjében. És nota bene, egy elsötétített akváriumban a vaklazacok mindig sokkal hatékonyabbak a táplálékszerzésben, mint látó társaik.
(Ui.: A fejbe besüllyedt, amorf szem természetesen éppen olyan csökevény szerv, mint pl. az emberi farok, vagy a korábban tárgyalt más példák.)
Menuet, A, Alunni, A, Joly, JS, Jeffrey, WR, Rétaux, S (2007) Expanded expression of Sonic Hedgehog in Astyanax cavefish: multiple consequences on forebrain development and evolution. Development 134: 845-855.
Jeffrey, WR (2005) Adaptive evolution of eye degeneration in the mexican blind cavefish. J. Hered. 96(3): 185-196. doi: 10.1093/jhered/esi028
Yamamoto, Y, Stock, DW, Jeffrey, WR (2004) Hedgehog signalling controls eye degeneration in blind cavefish. Nature 431: 844-847.




