



Az ecetmuslica széles körben használt kísérleti állat, még Thomas Hunt Morgan kezdte őket vizsgálni a laborjában 1907 -ben. Mivel jó páran követték a példáját, az évek alatt rengeteg ecetmuslica törzset alapítottak, amik a mai napig fennmaradtak. Nem kell bonyolult folyamatra gondolnunk, egy törzsalapítás során fognak pár ecetmuslicát, beteszik őket egy üvegfiolába, aminek az aljára valamilyen táptalajt helyeznek (például főtt krumplit vagy banánt), a száját pedig bedugják egy vattadugóval, aztán néhány hetente friss üvegbe rázzák át őket. Így a legyek csak egymás között szaporodhatnak, a többi ecetmuslicával semmilyen módon sem keverednek. Az évek alatt rengeteg ilyen törzset alapítottak, a legrégebbiek már több mint száz éve élnek fajtársaiktól elzárva (ha valakit érdekel, még mindig megvan a Thomas Hunt Morgan által 1910 -ben leírt, a w1 nevű, a világon elsőként megfigyelt mutációt hordozó törzs, pár dollárért elküldik postán is). Na de mi köze mindennek a genomokhoz? A hatvanas-hetvenes években jó páran elvégezték a kísérletet, hogy frissen, a vadonból befogott ecetmuslicákat kereszteztek a laboratóriumokban élő törzsekkel és egészen megmagyarázhatatlan jelenséget figyeltek meg (Pl: Engels és Preston 1979): Ha vadon élő nőstényeket kereszteztek laborhímekkel, akkor az utódokon semmi különleges sem látszott, egészségesnek tűntek, életképesnek. Viszont a fordított keresztezés során, amikor vadonból befogott hímeket kereszteztek labornőstényekkel, az utódok mindenféle mutációkat hordoztak, amik a szülőkben még nem voltak meg, sőt, a hőmérséklet emelésével a peték nagy részéből nem is kelt ki semmi (az első keresztezésből rakott peték 98-99% -ból kikelt a lárva 21, 25, 29 Celsius fokon is, a második keresztezés esetén 21 fokon a peték 96% -ból kelt ki lárva, 25 fokon csak a felükből, ám 29 fokon egy sem bizonyult életképesnek). A vad hímek vad nőstényekkel és a labornőstények laborhímekkel továbbra is egészséges, életképes utódokat hoztak létre. Meglepő eredmény, számításba véve, hogy a legrégebbi labortörzsek ekkor hetven éve szakadtak csak ki a vad ecetmuslicák közül.
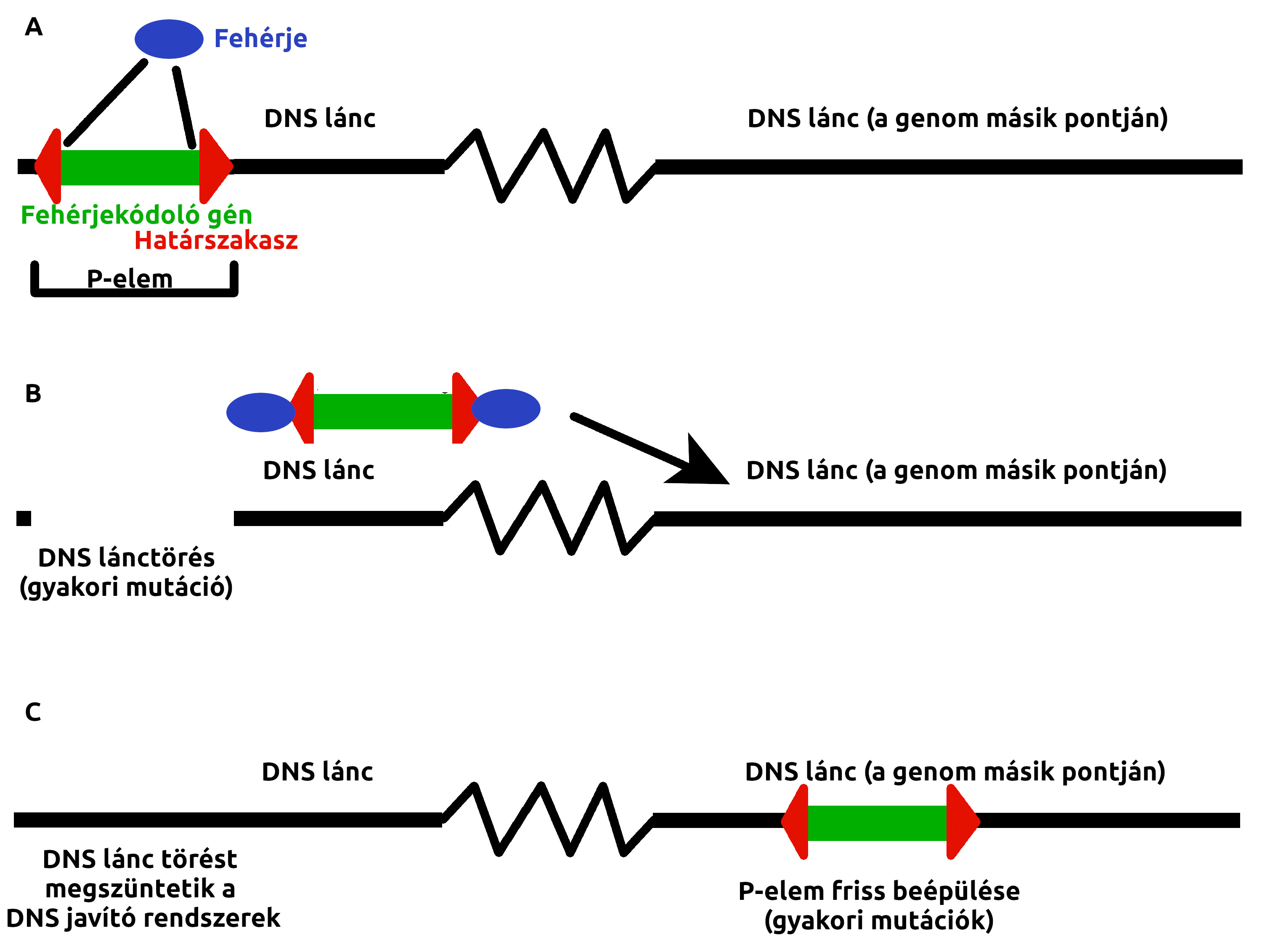
Ez a jelenség annak idején teljesen rejtélyesnek tűnt: Láthatóan valamilyen öröklődő jelleg okozza ezt az elharapózó terméketlenséget és a rengeteg mutációt, de az öröklődése semmilyen ismert szabályt sem követett. Évekkel később derítették fel az okát: A vad ecetmuslicák genomja egy mozgékony genetikai elemet tartalmazott, amit P-elemnek neveztek el. A mozgékony genetikai elem (vagy más néven transzpozon) nyilván nem annyira közismert fogalom, de nem is egyszerű körülírni. Nem vírus, nincs fertőző alakja, egyik egyed nem kapja el a másiktól, úgy terjed, hogy egy hordozó és egy nem hordozó egyed utódai mind hordozzák majd. Talán a legközelebb akkor jutunk a lényegéhez, ha úgy képzeljük el, mint fertőző genetikai információt. A genomi DNS egy szakasza, ami a világon semmiben sem különbözik a genom többi szakaszától, ugyanolyan DNS szál, ugyanúgy lemásolja a sejt osztódáskor. Ellenben egy ilyen P-elem háromezer bázispárnyi területét mindkét oldalon egy-egy jellegzetes bázissorrendű DNS szakasz határolja: CATGATGAAATAACATAAGGTGGTCCCGTCG és a másik oldalról CGACGGGACCACCTTATGTTATTTCATCATG. Így nem sok mindent mutat a két határszakasz, nincs is más szerepük, mint hogy a genomban felismerhetőek legyenek a bázissorrendjük alapján. A két határszakasz között található egyetlen gén, ami egyetlen fehérjét kódol (A ábra). Ez a fehérje egyetlen dolgot tesz: Megkeresi a genomban a P-elem két határszakaszát, a köztük lévő örökítőanyaggal együtt kivágja őket a genomból és egy másik, véletlenszerűen kiválasztott helyre illeszti be (B és C ábra). Ez a magyarázata a rengeteg megjelenő mutációnak, amikor a P-elem véletlenszerűen beül valahová a genomba, ott háromezer bázispárnyi friss DNS jelenik meg, ez könnyedén elront bármilyen gént. A másik oldalról pedig, amikor egy P-elem elmozdul a genomban addig elfoglalt helyéről, a helyváltoztatás időnként DNS szakaszok eltűnésével jár, nyilván egy gént könnyedén működésképtelenné tesz, ha eltűnik belőle néhány tucat bázispárnyi DNS. A sok frissen helyet változtató P-elem sok gént ront el, minél többet, annál valószínűbb, hogy életképtelenné teszi az egyedet. A P-elem tökéletesen illeszkedik Richard Dawkins önző gén fogalmába: Kizárólag önmagáért létezik, a határszakaszok azért vannak, hogy a P-elem által kódolt fehérje felismerje, melyik szakaszt kell kivágni a genomból, a P-elem egyetlen génje pedig a saját magát a genomban áthelyező fehérjét kódolja. Más működése nem ismert, más örökítőanyagot nem tartalmaz, helyet változtat a genomban, saját magát szaporítja. Nincs saját teste, még egy fehérjeburka sem, mint egy vírusnak, a genomon kívül nem létezik, teljesen életképtelen, semmilyen életműködést sem mutat, egyszerűen egy egységnyi önmagát másoló, fertőző genetikai információ.
De miért csak a labornőstény vad hímmel keresztezésekor figyelhető meg ez a jelenség? A P-elemek sem védettek a mutációktól, időnként ezek is elromlanak, a genombeli mozgásuk során mindenféle P-elem töredékek keletkeznek, amik különbözőképpen váltak működésképtelenné. Például ha egy P-elem félig működőképes fehérjét termel, ami megkeresi a genomban a határszakaszokat, hozzájuk kötődik, de nem képes kivágni a helyéről a P-elemet, akkor ez bizony akadályozza az esetleges működőképes fehérjéket is, mert azok nem férnek oda tőle a határszakaszokhoz. Egy ilyen mutáció nyomán egy P-elemből máris a többi hasonló elem mozgását gátló gén alakult ki, ha egy sejtmagban ilyen defektes P-elemek is előfordulnak, akkor az ezekről átíródó hibás fehérje az ép P-elemek mozgását is akadályozza, a jószág genomjában máris alig mozognak a P-elemek, így védett az így létrejövő mutációktól is. A vadon élő muslicák genomja mind hasonló gátlóelemeket tartalmaz, így lehetnek életképesek és egészségesek annak ellenére, hogy a genomjukban P-elemeket is tartalmaznak. Ezért növekszik meg ugrásszerűen a mutációk száma, ha labornőstényeket keresztezünk vadonból befogott hímekkel: A petében nem termelődnek a P-elem helyváltoztatását gátló fehérjék, az apai genom viszont tartalmaz P-elemeket, amik így teljesen szabadon mozognak. A mozgékony genetikai elem megszelídítése viszonylag gyorsan lejátszódik, kipróbálták hogy mi történik (Kidwell és mtsai. 1988), ha labortörzsekbe frissen juttatnak be egy ép, működőképes P-elemet: Nem meglepő módon egy ilyen esemény nyomán hirtelen elterjed a törzsben a P-elem, hamarosan minden egyed hordoz belőle jó párat a genomjában, majd 20-40 nemzedéknyi idő alatt megjelennek a P-elem helyváltoztatását gátló hibás P-elemek, onnantól kezdve az egyedek védettek a káros hatásoktól. Vagyis egy ecetmuslica törzs viszonylag hamar megszelídít egy ilyen bejutó mozgékony genetikai elemet, onnantól az egyedek megint teljesen életképesek, termékenyek, nem keletkeznek bennük fölös számban mutációk a P-elem helyváltoztatása nyomán.
Kézenfekvő kérdés, hogy de akkor miért nincsenek meg ezek a labortörzsekben is? A vadonban befogott minden légytörzs tartalmaz P-elemeket, Indokínától Patagóniáig, miért nincs a labortörzsekben? A válasz megtalálásában éppen a genetika úttörői segítettek: A kezdetektől alapítottak új törzseket, ezek mind egy-egy apró ablakként működnek az alapításkori ecetmuslica genomra, hiszen csak egymás között szaporodva külső hatásoktól mentesen élnek. Kidwell 1983 -ban egyszerű kísérletet talált ki: Összegyűjtött egy csomó régi törzset és ellenőrizte, hogy vajon P-elemeket tartalmaznak -e? Az eredmény meg is magyarázza a jelenséget: Az 1920 -as években alapított négy törzsből egy sem tartalmazott P-elmeket, ugyanígy ez a mozgékony genetikai elem hiányzott a harmincas és a negyvenes években alapított tizennégy illetve hét törzsből is. Azonban az ötvenes években alapított harminckilenc törzsből már négy P-elemeket is hordozott (10,2%), a hatvanas években alapított hetvenkilenc törzsből huszonegyben talált P-elemeket (26,6%), a hetvenes években alapított, vagyis a vadonból befogott kétszázötvenhét törzsből kétszázhuszonöt P-elemeket hordozott (87,6%). A később alapított törzsek közül már mindegyik P-elemeket is hordoz, ez az állapot jellemző mind a mai napig is. A magyarázat ez alapján egyszerű: A labortörzsek genomjában azért nem találni P-elemeket, mert akkoriban ezek a vadon élő ecetmuslicák genomjába se jutottak be, egyszerűen ezek a populációk pillanatképek a korabeli vadon élő ecetmuslicák genomjáról. Valamikor a negyvenes években, amíg mi éppen világháborút vívtunk bejutott ennek a vadon élő fajnak a genomjába a P-elem, egy sajátos öncélú genomi élősködő, szépen elkezdte meghódítani a világot. Amíg mi forradalmat csináltunk, a P-elem tovább hódított, mire felépítettük az érett Kádár-rendszert, már a világon mindenfelé élő ecetmuslicák között rég többségbe kerültek a P-elemet hordozó egyedek, a rendszerváltás idejére pedig gyakorlatilag nem maradt a Földön P-elem mentes ecetmuslica, ez a sajátos gén Afrikától Brazíliáig mindenhol elterjedt röpke negyven év alatt. Egy vadon élő állatfaj genomját teljes egészében felforgatta egy újonnan bejutott mozgékony genetikai elem emberi ésszel könnyedén belátható idő alatt. És ami a laikus olvasónak a leghihetetlenebb lehet: Észrevétlenül. Ez a nagy horderejű változás senkinek sem tűnt föl, nem borított föl életközösségeket, nem okozott semmilyen környezeti kárt. A vadonatúj gént az ecetmuslica genom különösebb nehézségek nélkül megszelídítette. Mostanra láthatóan nyugvó állapotba került, ritkán változtat helyet az ecetmuslica genomban, különböző gátlóelemek tartják így.
Mennyire kivételes példa ez az eset? Vajon valami kozmikus véletlen, amit pont szerencsésen megfigyeltünk, de nem várható soha többé, vagy gyakori? Meglepően gyakori. Például a P-elem nem állt meg az ecetmuslicánál, egy nagyon közeli rokon faj, a Drosophila simulans floridai és dél-afrikai populációiból is kimutatták a P-elemek jelenlétét 2010 -ben és 2012 -ben. Floridában egyedenként átlagosan 0,4 P-elemet tartalmaztak a genomok, Afrikában 29 -et, jelezve, hogy Floridában még éppen csak megindult a P-elem térhódítása, míg Dél-Afrikában már egy egyenletesen elterjedt fertőzésről beszélhetünk (Kofler és mtsai. 2015). Ez az eredmény azért is beszédes, mert ebben a fajban egy 2001 -ben Afrikában végzett felmérés nyomát sem találta P-elemeknek, és az 1998 előtt alapított D. simulans törzsekben sem találtak P-elemeket. Minden jel szerint ez a mozgékony genetikai elem megint átlépte a fajhatárt és éppen egy új muslicafajt hódít meg jelenleg is, amíg e sorokat írom. Nem meglepő módon ez is teljesen észrevétlenül zajlik, egy ilyen nagymértékű genomi változás semmilyen környezeti következménnyel sem jár.
Ezen kívül megvizsgálhatjuk az ismert genomokat, például az emberi genom majdnem felét teszik ki a különböző megszelídített mozgékony genetikai elemek, eddig minden megismert genomban rengeteg ilyen egykor mozgékony elem roncsai terpeszkednek. Minden jel szerint a földi élőlények körében ezek a különböző fertőző genetikai információcsomagok általánosan elterjedtek, időnként megugorják a fajhatárokat, olyankor meghódítják egy új faj genomját, aztán hamar megszelídülnek.
Engels, W. R., & Preston, C. R. (1979). Hybrid dysgenesis in Drosophila melanogaster: the biology of female and male sterility. Genetics, 92(1), 161-174.
Castro, J. P., & Carareto, C. M. (2004). Drosophila melanogaster P transposable elements: mechanisms of transposition and regulation. Genetica, 121(2), 107-118.
Kidwell, M. G., Kimura, K., & Black, D. M. (1988). Evolution of hybrid dysgenesis potential following P element contamination in Drosophila melanogaster. Genetics, 119(4), 815-828.
Kidwell, M. G. (1983). Evolution of hybrid dysgenesis determinants in Drosophila melanogaster. Proceedings of the National Academy of Sciences, 80(6), 1655-1659.
Kofler, R., Hill, T., Nolte, V., Betancourt, A. J., & Schlötterer, C. (2015). The recent invasion of natural Drosophila simulans populations by the P-element. Proceedings of the National Academy of Sciences, 112(21), 6659-6663.




