 Hogy mindjárt a közepébe vágjak, mert én őszinte vagyok, még ha te, kedves olvasó, prűd is: péniszekről lesz szó, pontosabban falloszokról, sőt, hogy ne verjük ki minden tartalom-szűrő biztosítékát, phallusokról. Mitöbb, nem is róluk, inkább a hiányukról.
Hogy mindjárt a közepébe vágjak, mert én őszinte vagyok, még ha te, kedves olvasó, prűd is: péniszekről lesz szó, pontosabban falloszokról, sőt, hogy ne verjük ki minden tartalom-szűrő biztosítékát, phallusokról. Mitöbb, nem is róluk, inkább a hiányukról.
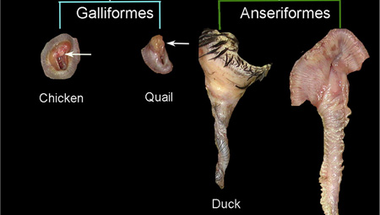
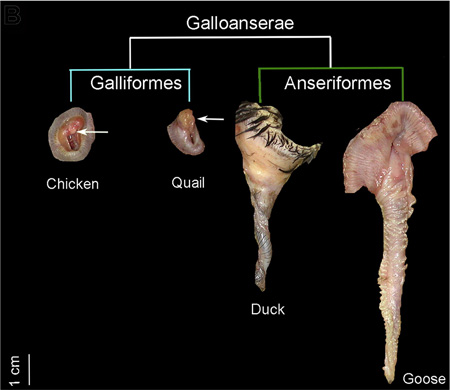
Ezzel a felvezetővel talán már mindenkit kellően összezavartam, szóval kevésbé kódolt formában is legyen mindez megfogalmazva: mint néhány kacsás posztban korábban már írtunk róla, egyes madárfajok hímjeinek kifejezetten impresszív hímtagja van, ami azért is szembetűnő mert számos más fajnál (pl. a házityúk) egyáltalán nincs ilyesmi, párzáskor a hímek a kloákanyílásukat szorítják a nőstények kloákájára, hogy átjuttassák az ivarsejtjeiket.
A nemiszerv ilyenfajta nem-meglétét a szakirodalom "phallus non-protrudens"-ként ismeri (ami számos összehasonlító anatómia tankönyv egyik legszórakoztatóbb kifejezése is) és egy fejlődésbiológust, különösen a kacsák esetét észben tartva, természetesen könnyen lázba hoz a kérdés, hogy mi okozza a nagy különbségeket a csoporton belül.
A napokban megjelent Current Biology cikk egy kicsit segíti a tisztánlátást. Ugyanis kiderült, hogy tulajdonképpen a fejlődés korai szakaszaiban kakasokban is megjelenik a phallus, csak növekedése a gácsorkéhoz képest lelassul, később pedig egyenesen elsatnyul.
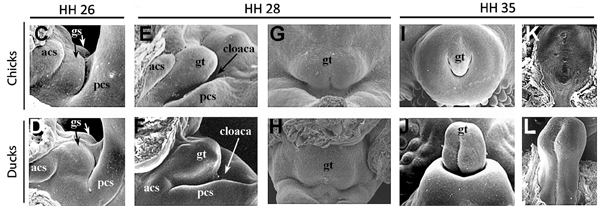
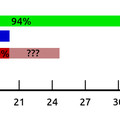
A szó klasszikus értelmében vett "csökevény szervről" van hát szó, amelynek fejlődési dinamikája sokban hasonlít pl. az emberi farokéra. S hogy az analógia még jobb legyen a jelek szerint ugyanaz a folyamat, a programozott sejthalál, ami azért felelős, hogy a phallus sejtjei elhalnak (az alábbi ábra felső sorában különböző fajok fejlődő phallusai láthatók, és a piros pöttyök elhaló sejteket jelölnek - látható, hogy a csirke és a hozzá hasonló fürj esetében ez igen intenzív folyamat, míg a kitüremkedő nemiszervvel rendelkező fajok esetében koránt sincs ennyi elhaló sejt).
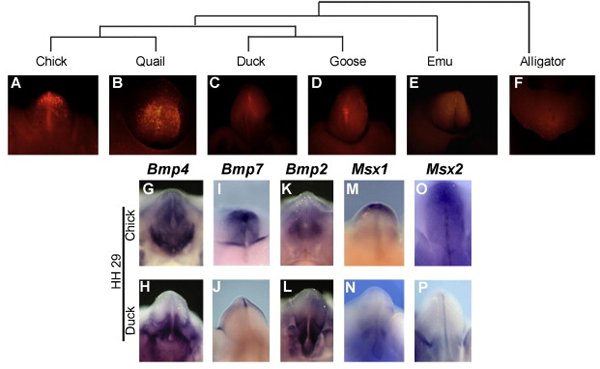 Ebből persze adódik is a kérdés, hogy vajon mi okozhatja a sejtek elhalását? Szerencsére, itt máris adódik egy jó jelölt, a BMP-jelátviteli útvonal, amely számos más kontextusban (például a lábujjak közti szövetrészekben) felelős az apoptózisért. És valóban, ebben az esetben is, lényeges különbség egy csirke és egy kacsa között, hogy a Bmp ligandumokat kódoló gének, illetve a BMP-jelátvitel célgénjeiként működő Msx gének expressziója megnő a fejlődő kakasok phallusában, jelezve, hogy aktív BMP-szignalizáció folyik a szövetrészben.
Ebből persze adódik is a kérdés, hogy vajon mi okozhatja a sejtek elhalását? Szerencsére, itt máris adódik egy jó jelölt, a BMP-jelátviteli útvonal, amely számos más kontextusban (például a lábujjak közti szövetrészekben) felelős az apoptózisért. És valóban, ebben az esetben is, lényeges különbség egy csirke és egy kacsa között, hogy a Bmp ligandumokat kódoló gének, illetve a BMP-jelátvitel célgénjeiként működő Msx gének expressziója megnő a fejlődő kakasok phallusában, jelezve, hogy aktív BMP-szignalizáció folyik a szövetrészben.
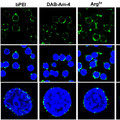
De állhat-e ez a korreláció egyben egy ok-okozati összefüggés mögött? Ennek megvélaszolására egy BMP-jelátvitel antagonistát, Noggint tartalmazó golyócskákat ültettek a kakasembriók fejlődő phallusába és figyelték a hatást.
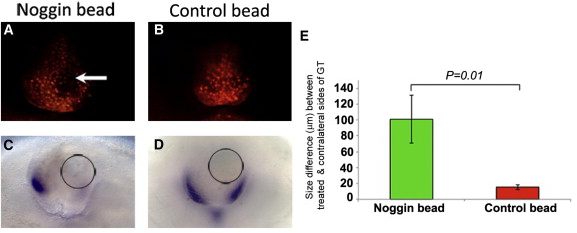
A különbség leglátványosabb része, hogy a sejthalál a golyó környékén szinte megszűnik, ahogy az Msx expresszió is (vagyis tényleg nincs aktív BMP jelátvitel errefele). És a sejthalál elmaradása látványos hatással bír: az a phallus-oldal, ahol a golyó volt, lényegesen nagyobb lesz, mint a másik oldal. (Természetesen, egy kontroll golyóval mindez a változás elmarad.)
Mindez tehát egyfajta mechanisztikus magyarázatot ad a jelenségre, ugyanakkor marad a nagy kérdés: miért? Evolúciósan miért nincs ezeknek a fajoknak ugyanolyan nemiszerve, mint például a kacsáknak. A válasz nem egyértelmű, pontosabban csak találgatni tudunk. Lehet, hogy a kloakák összenyomása egy koordináltabb folyamat, mint a klasszikus penetráció, így a nőstények is jobban kontrollálni tudják, hogy kivel szűrik össze a levet - magyarán egyfajta szexuális szelekció eredménye, amit látunk. Egy másik lehetőség, hogy a BMP-jelátvitel megváltozása csak avalamilyen más, evolúciósan fontos jelenség mellékzöngéje, és mivel ez a bizonyos obskurus tulajdonságra erős szelekció volt, a csökevényesedett phallus is megmaradt. De az igazság az, hogy mindkét hipotézisre számos ellenérv is létezik, így ennek az evolúciós oknak a dekódolása még az utókorra vár.
Herrera AM, Shuster SG, Perriton CL, Cohn MJ (2013) Developmental Basis of Phallus Reduction during Bird Evolution. Curr Biol 23(12): 1065-1074.