 Richard Lenskit nyilván senkinek sem kell bemutatnom, lebilincselően izgalmas kísérletei (1, 2, 3) elég sok adalékot szolgáltattak az evolúció megismeréséhez. Kétségkívül leghíresebb munkája az évtizedek óta folyó E. coli evolúciós kísérlete, ahol egy E. coli törzset többfelé osztott, majd szőlőcukorban szegény, citromsavban gazdag táptalajra helyezte őket és figyelte, mi történik velük. Az E. coli egyik jellemzője ugyanis, hogy csak anaerob körülmények között képes citromsavat hasznosítani, oxigén jelenlétében a citromsav nem jut át a sejtmembránon, így nem is képes táplálékként felhasználni a sejt. A mikrobiológiai gyakorlatban ez alapján azonosítják E. coliként. Lenski eredeti feltételezése az volt, hogy mivel az E. coli anaerob körülmények között képe citromsavat is hasznosítani, megfelelő körülmények között előbb-utóbb az evolúció során kialakul az oxigén jelenlétében való citromsavfogyasztás képessége is. Nem csalódott, röpke 31000 nemzedék után a tizenkét törzs egyikében meg is jelentek citromsavat is hasznosító sejtek, ezeket nevezte el Cit+ -nak.
Richard Lenskit nyilván senkinek sem kell bemutatnom, lebilincselően izgalmas kísérletei (1, 2, 3) elég sok adalékot szolgáltattak az evolúció megismeréséhez. Kétségkívül leghíresebb munkája az évtizedek óta folyó E. coli evolúciós kísérlete, ahol egy E. coli törzset többfelé osztott, majd szőlőcukorban szegény, citromsavban gazdag táptalajra helyezte őket és figyelte, mi történik velük. Az E. coli egyik jellemzője ugyanis, hogy csak anaerob körülmények között képes citromsavat hasznosítani, oxigén jelenlétében a citromsav nem jut át a sejtmembránon, így nem is képes táplálékként felhasználni a sejt. A mikrobiológiai gyakorlatban ez alapján azonosítják E. coliként. Lenski eredeti feltételezése az volt, hogy mivel az E. coli anaerob körülmények között képe citromsavat is hasznosítani, megfelelő körülmények között előbb-utóbb az evolúció során kialakul az oxigén jelenlétében való citromsavfogyasztás képessége is. Nem csalódott, röpke 31000 nemzedék után a tizenkét törzs egyikében meg is jelentek citromsavat is hasznosító sejtek, ezeket nevezte el Cit+ -nak.
Mivel szabályos időközönként mind a tizenkét törzsből fagyasztottak le mintákat, mintegy újra lejátszhatták az evolúciójukat, azt tapasztalták, hogy a Cit+ törzsben a késői nemzedékekben nagy gyakorisággal jelentek meg újonnan Cit+ egyedek, míg a koraiakban egyáltalán nem. Ezt úgy magyarázták, hogy valamilyen mutáció rögzült ebben a törzsben, amely önmagában nem elég a citromsav felhasználásához, de ezen a genetikai háttéren már egyetlen mutáció is elég ennek a jellegnek a megjelenéséhez. De mi is a Cit+ törzs citromsav hasznosításának az alapja? Végre tavaly szeptemberben leközölték ezt is, úgyhogy nem kell tovább találgatnunk.
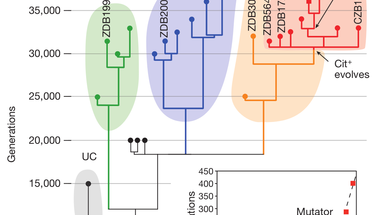
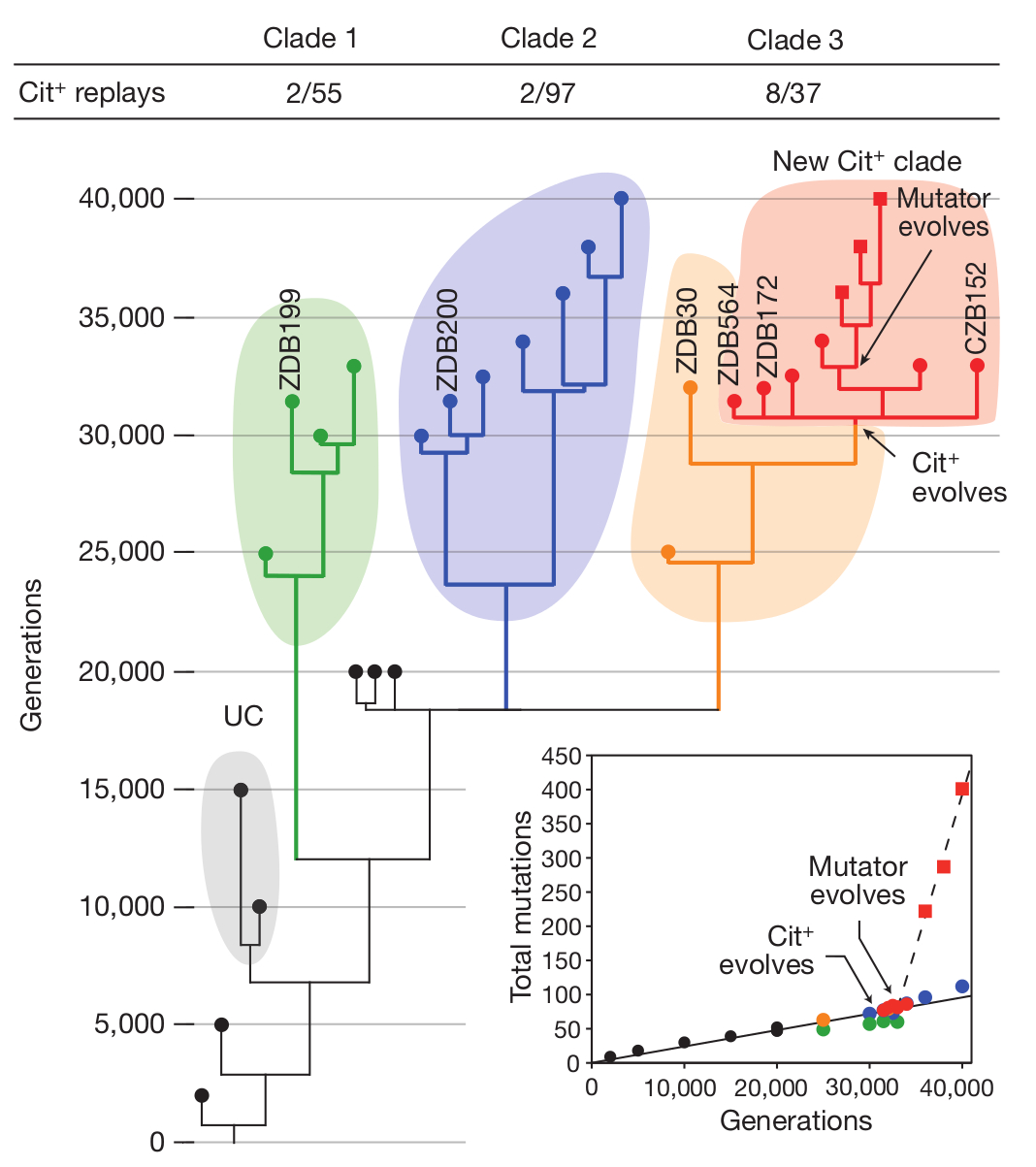
Különböző időpontokból vett huszonkilenc klón genomját szekvenálták újra, így lépésről-lépésre fölrajzolhatták, hogyan alakult ki a Cit+ törzs, ezek közt hat Cit+ és három Cit- is volt, amelyekről már korábban kimutatták, hogy belőlük nagy gyakorisággal alakulnak ki Cit+ törzsek. Meglepő módon azt találták, hogy a populáció egész végig polimorf volt, azaz több különböző törzs élt egymás mellett, ám ezek közül csak az egyikben alakult ki a citromsav felhasználásának képessége, ez látható az első ábrán. Az egyik, (UC) tizenötezer nemzedéken át bírta, de végül kihalt a populációból. A C1, C2 és C3 törzsek a kísérlet folyamán végig egymás mellett léteztek, de csak a C3 törzs egyedei közt jelentek meg citromsavat is felhasználók. Hogy miért léteztek ilyen sokáig egymás mellett a különböző törzsek, két magyarázat lehetséges: Az első, hogy a kultúrában külön niche -eket töltöttek be, az egyik az egyik anyagcseretermék hasznosítására szakosodott, a másik egy másikéra. A másik magyarázat, hogy egyikben sem történt olyan mutáció, ami döntő előnyhöz juttatta volna a többiekkel szemben, így nem tudta azokat kiszorítani. (A C3 törzsben jelent meg egy pontmutáció a mutS génben, aminek a következtében osztódáskor sokkal több mutáció történt a sejtekben, meglepő módon ez a mutátor fenotípus a Cit+ fenotípus után alakult ki. )
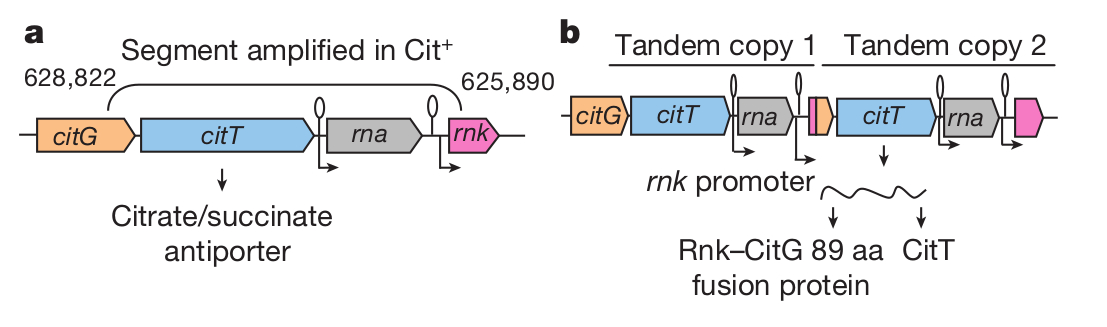
A Cit+ jelleg három lépésben alakult ki. Először megjelent egy olyan genetikai háttér, amelyen nagy gyakorisággal alakultak ki Cit+ egyedek, majd megjelent maga a Cit+ fenotípus, ezek a sejtek még alacsony hatékonysággal hasznosították a citromsavat, majd a hatékonyságuk jelentősen javult, ez tette lehetővé a Cit+ populáció dominánssá válását. Mindegyik Cit+ törzsben egynél több példányban található meg a cit operon, egészen pontosan az rna (ami egy RNáz I33 nevű fehérjét kódol) és a citT (ami egy citromsav/szukkcinát antiporter fehérjét kódol, ez juttatja át a sejtmembránon a citromsavat anaerob körülmények között) gének duplikálódtak. Ez a duplikáció nem található meg egyik Cit- törzsben sem, így feltételezhetően szükséges a citromsav emésztéséhez. Ahogy a második ábrán látható, ez a genom átrendeződés azzal járt, hogy a duplikálódott szakasz két végén található gének egy szakasza is duplikálódott, így az új gén az rnk (nukleozid difoszfát kinár szabályozófehérjét kódol), a citG (trifoszforibozil-defoszfo-CoA szintáz enzimet kódol) gének egy-egy rövidebb szakaszát tartalmazza, valamint a citT gén egészét, így a róla átíródó fehérje az RNK fehérje N-terminális végével kezdődik, utána a CITG fehérje C-terminális végével folytatódik, amit az rnk gén promótere hajt meg. Ez azért fontos, mert az rnk gén oxigén jelenlétében is átíródik, így mivel a citT gén eredetileg a citG -vel monocisztronos, az rnk promóter ezután az újonnan duplikálódott citT -gént is meghajtja, így az eredetileg csak oxigén hiányában termelődő CitT fehérje ez után oxigén jelenlétében is kifejeződik. Ez a kiméra gén egy Cit- törzsbe juttatva egészen gyengén Cit+ fenotípust eredményez, ami annyira rosszul hasznosítja a citromsavat, hogy a szőlőcukor elfogyása után hatvan órával mértek valami gyenge növekedést. Mivel az eredeti törzset naponta friss táptalajra helyezték, ez szinte semmi előnyt sem jelentett az első Cit+ egyedeknek, a mérések szerint ez a törzs 1% -nyi kompetitív előnyt élvezett a szülői törzshöz képest.
A későbbi nemzedékekből (~33k) származó Cit+ egyedek genomjában ez a duplikáció újra duplikálódott, a citromsav felhasználásának hatékonyabbá válásával párhuzamosan nőtt a citT kópiaszáma, a későbbi egyedek kilenc példányt is hordoztak az eredeti duplikációból. Ez tette lehetővé a citromsavat felhasználó populáció elszaporodását. Meglepő módon később (~35k) a genomban található rnk-citT modulok száma újra lecsökkent, miközben a citromsav felhasználás hatékonysága nőtt, ezzel egy időben pontmutációk rögzültek magában a citT génben, a gltA nevű citromsav-szintáz enzimet kódoló génben és az aceA izocitromsav liáz enzimet kódoló génben, vélhetőleg ezek felelősek a megfigyelt hatékonyságnövekedésért, de ezt a cikkben nem részletezik, mondjuk nem is túl érdekes.
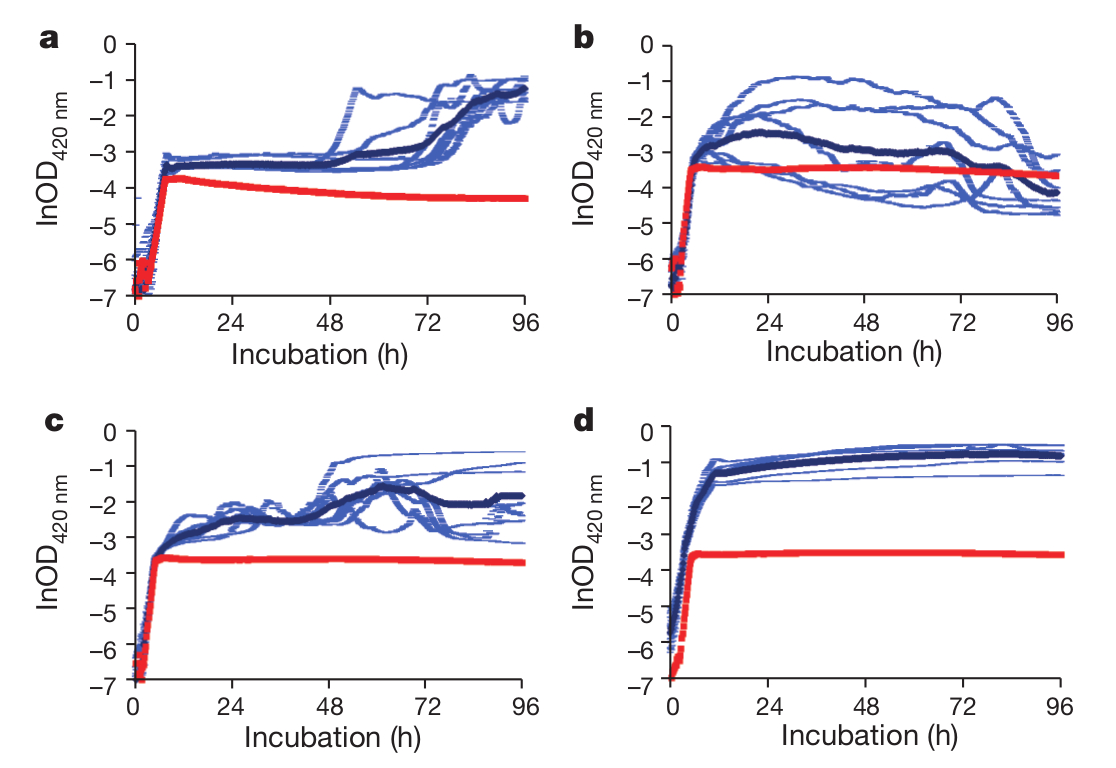
Ez eddig szép és jó, de a legfőbb kérdés továbbra is érintetlen maradt: Ha ennyire egyszerű, egy duplikáció kellett csak a Cit+ fenotípus megjelenéséhez, miért tartott ilyen sokáig? A válasz annyi, hogy ezek a mutációk csak korábbi mutációk után alakítják ki a Cit+ fenotípust. Kipróbálták ugyanis, hogy mi történik, ha egy rnk-ciT modult juttatnak a nulladik generáció genomjába? Meglepő módon ez sokadik próbálkozásra sem sikerült, valószínűleg azért, mert egyetlen rnk-citT modul még nagyon gyenge Cit+ fenotípust eredményez. Fogták és ezt a modult beépítették egy nagy kópiaszámú pUC plazmidba, így juttatták be a sejtekbe, ez a több száz kópiában jelen lévő rnk-citT modul már elég volt egy tisztességes Cit+ fenotípushoz, ezek a sejtek már felhasználták a citromsavat, ám meglepő módon a C1 és C2 törzs még így is rosszabbul hasznosította a citromsavat, mint a C3 törzs, ez látható a harmadik ábrán. Ebből egyértelműen látszik, hogy a C3 törzsben már a Cit+ jelleg megjelenése előtt rögzültek olyan mutációk, amik hatékonyabbá tették a sok ezer nemzedék múlva megjelenő citromsavhasznosítást. Az "a" a szülői törzs, a "b" a C1, a "c" a C2, a "d" a C3 törzs. (A képen a piros a szülői törzs kísérleteinek az átlaga, sötétkékkel a plazmidot hordozó törzs kísérleteinek az átlaga látható, világoskékkel pedig az egyes plazmidot hordozó törzsekkel végzett kísérleteket mutatják be. A vízszintes tengely az idő, a függőleges tengely a kultúra optikai denzitása.) Sajnos nem sikerült egyértelműen megmutatniuk mely mutáció(k) felelősek ezért a jelenségért.
Felmerült még egy kérdés, hogy ez a mutáció csak az rnk promóterrel működik -e vagy bármilyen promóter jó, ami aerob körülmények között is meghajtja a citT gént? Egy Cit- törzsben újra hagyták kialakulni a Cit+ fenotípust, aztán megvizsgálták, hogy milyen mutációk okozták. Hét esetben a fent leírthoz hasonló rnk-citT modul alakult ki, hat esetben egy IS3 ugráló genetikai elem ült a citT gén elé, amely egy általános promótert hordoz, két esetben nagy duplikációk történtek, amelyek az egész operont érintik, egy mutánsban pedig egy óriási inverzió történt, amely a genom távoli pontjára, egy másik gén promóterének szabályozása alá helyezi a citT gént egy másik mutánsban pedig egy deléció történt a citT gén előtt, ami egy teljesen új promótert hozott létre. Ebből is látszik, hogy ez nem valami különlegesen ritka mutáció, a megfelelő genetikai háttéren többféleképpen létrejöhetnek Cit+ egyedek.
Azért kicsit csalódott voltam a cikk olvastán, mivel a legfőbb kérdésre most sem kaptam választ, hogy vajon mi(k) azok a mutáció(k), amely(ek) önmagukban nem elégségesek a citromsav aerob körülmények közti felhasználásához, de valamiért ilyen mértékben módosítják a citT gén mutációinak a hatását. Remélem előbb-utóbb ez is kiderül majd!
Blount, Z. D., J. E. Barrick, C. J. Davidson, and R. E. Lenski. 2012. Genomic analysis of a key innovation in an experimental Escherichia coli population. Nature 489:513-518.