A biológiában a nagyon hasonló formák (legyenek azok akár testrészek, akár genetikai hálózatok, akár sejtalkotók) jelenléte két szervezetben nagyon jó eséllyel közös eredetre utalnak.
A biológiában a nagyon hasonló formák (legyenek azok akár testrészek, akár genetikai hálózatok, akár sejtalkotók) jelenléte két szervezetben nagyon jó eséllyel közös eredetre utalnak.
Megvan ennek a maga logikája, természetesen: hiszen ha A forma jelen volt a közös ősben, akkor a két faj szétválása után a két vonalban szabadon módosulhatott és kialakíthatta a különböző, ugyanakkor mégiscsak hasonló A' és A" formákat.
A levezetés tetszetős és többnyire igaz is, de nem mindig. A nagy és fontos különbség az ún. konvergens evolúció miatt van. Ez az a jelenség lenne, amikor egy biológiai probléma megoldása annyira optimális, hogy különböző kiindulási pontokból is nagyon hasonló fenotípusok jönnek létre a természetes szelekciónak köszönhetően (gondoljunk csak egy egyaránt vízi életre berendezkedett Ichtyosaurus és delfin közötti hasonlóságra, vagy akár az echolokációt használó cetek és denevérek hallásában szerepet játszó fehérjék párhuzamos evolúciójára).
Most a konvergens evolúció példatára egy új, igen látványos darabbal gazdagodott, mégpedig a különböző fajok harántcsíkolt izmainak esetével.
A szervezet legfőbb mozgatórugóit jelentő harántcsíkolt izmoknak igen jellegzetes periódikus felépítése van, ami már viszonylag kis nagyítású mikroszkópon is kivehető és amelynek okát egy valódi, molekuláris ismétlődésben kereshetjük.
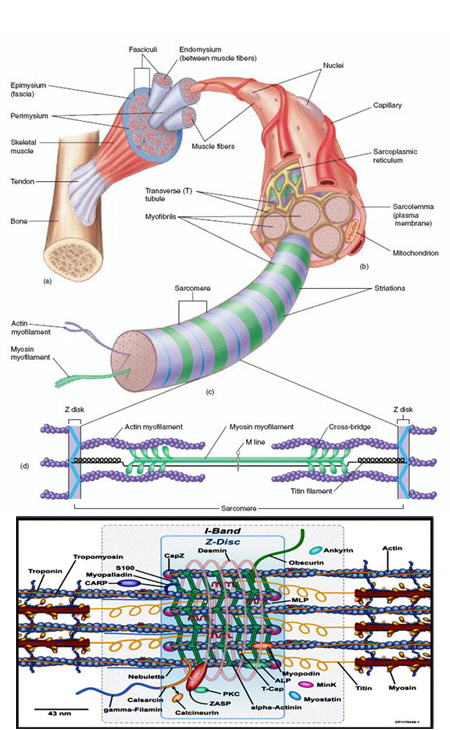 Ha egy tipikus gerinces harántcsíkolt izmát megnézzük, akkor hamar kivehető, hogy annak alapegysége az ún. izomrost, ami egy többsejtmagvú óriássejtként fogható fel.
Ha egy tipikus gerinces harántcsíkolt izmát megnézzük, akkor hamar kivehető, hogy annak alapegysége az ún. izomrost, ami egy többsejtmagvú óriássejtként fogható fel.
A rostokban miofibrillumokat lelünk, ezek pedig a mozgáshoz elengedhetetlen két fehérje, az aktin és a miozin rendezett elhelyezkedésével keletkeznek.
A rendezettség alapegysége a szarkoméra, amely két oldalán rögzített aktin polimereket tartalmaz (ezek a láncszerű sturktúták kis, gömbszerű, vagy szaknyelven globuláris fehérje alegységekből jönnek létre), amelyek közte található egy miozinköteg.
A miozinok motorfehérjékként működnek, amelyek a sejt energiáját biztosító ATP felhasználásával, megfelelő jel esetén elkezdenek végiglépkedni az aktinláncokon. Ez értelemszerűen a szakoméra rövidüléséhez vezet. Természetesen egyetlen szarkoméra csak nagyon kis rövidülést képes elérni még maximálisan összehúzódott állapotában is, de ha az egymás után elhelyezkedő sok-sok szarkoméra egyszerre húzódik össze, azzal már számottevő rövidülés érhető el.
Az aktin és miozin mellett még egy sereg járulékos molekula játszik szerepet a folyamatban (pl. a pihenő állapotban az aktin-miozin kötési felületet elrejtő troponin és tropomiozin, az szarkoméra visszanyúlásáért felelős, molekuláris rugóként működő titin), ezek közül mindenképpen említést érdemelnek azok, amelyek azt a vázat, az elektromikroszkópos képe alapján Z-vonalnak nevezett struktúrát alkotják, ahova az aktin láncok kötnek.
Érdekes módon rovarokban, ahol alapvetően nagyonon hasonló az izom felépítése, számos olyan fehérje vesz részt az izmok felépítésében, amely nem is fordul elő gerincesekben illetve vice versa. Vagyis az izmok, nagyon hasonló struktúrájuk ellenére evolúciós viszonylatban változóképes molekuláris alpokra építleznek. De még ebben a változékony világban is sikerült négy proto-Z-vonal komponenst azonosítani, olyan fehérjéket, amelyek minden kétoldali szimmetriájú állatban fellelhetőek. Ezek az α-actinin, az MLP (Muscle LIM Protein) és a Zasp.
Ezzel a tudással felvértezve, immár egy igen érdekes evolúciós kérdést lehet feltenni: a csalánozók közé tartozó egyes hidraafajokban (pl. Clytia haemispherica) jelen levő, mikroszkóposan harántcsíkolt izomnak tekinthető struktúrák mennyiben felelnek meg molekulárisan a gerincesek, vagy ízeltlábúak harántcsikolt izmának?
A válasz az, hogy csak igen mérsékelten. Ugyan ezeknek az állatoknak az izmai kifejezik a harántcsíkolt izmokra jellemző miozin-típus csalánozó ekvivalensét (Ch ST myhc-b), a geomjukban fellelhető MLP és zasp/ldb3 ortológok pont nem ezekben a sejtekben fejeződnek ki (az α-actinin alacsony szinten minden sejtben kifejeződik).
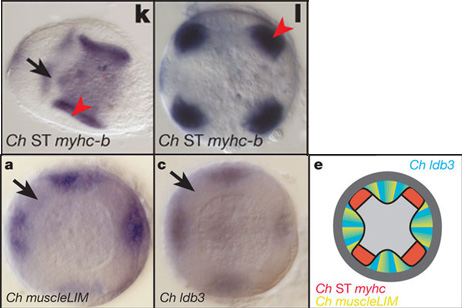
Vagyis a nagy hasonlóság ellenére, a csalánozók teljesen más molekuláris komponensekből hozták össze a maguk, "szarkomérákból" álló harántcsíkolt izmát. Hogy pontosan miből, az még a jövő zenéje, de érdekes látni, hogy egy ilyen komplex molekuláris gépzet is létrejöhet több módon is.
Hejnol A (2012) Evolutionary biology: Muscle's dual origins. Nature 487:181-2.
Steinmetz PR, Kraus JE, Larroux C, Hammel JU, Amon-Hassenzahl A, et al. (2012) Independent evolution of striated muscles in cnidarians and bilaterians. Nature 487:231-4.