Az állatvilágban több a miénktől eltérő rendszer létezik a nemmeghatározásra, ezek egyike a mézelő méh által alkalmazott. Az anya életében egyszer párzik, a begyűjtött spermiumokat egész életében a testében tárolja, amikor petét rak, eldöntheti, hogy megtermékenyíti, vagy nem. A megtermékenyítetlen petékből haploid herék (hímek) kelnek ki, a megtermékenyített petékből lesznek a diploid anyák és dolgozók. Őköztük annyi a különbség, hogyha egy lárva a fejlődése során nagy mennyiségben kap a méhek garatmirigyének hormontartalmú váladékából, akkor anyává fejlődik, ha nem kap ebből eleget, dolgozóvá. Csak az anyák termékenyek, ha egy család elveszíti az anyát, akkor a dolgozók egyike esetleg elkezdhet megtermékenyítetlen petéket rakni (ő ugye sohasem párzott), de attól a család kihal, így ez általában zsákutca, a család génjei élhetnek esetleg tovább, ha a lerakott petékből kikelő herék párosodnak egy anyával, de nyilván egy méhészen ez nem segít.
Az állatvilágban több a miénktől eltérő rendszer létezik a nemmeghatározásra, ezek egyike a mézelő méh által alkalmazott. Az anya életében egyszer párzik, a begyűjtött spermiumokat egész életében a testében tárolja, amikor petét rak, eldöntheti, hogy megtermékenyíti, vagy nem. A megtermékenyítetlen petékből haploid herék (hímek) kelnek ki, a megtermékenyített petékből lesznek a diploid anyák és dolgozók. Őköztük annyi a különbség, hogyha egy lárva a fejlődése során nagy mennyiségben kap a méhek garatmirigyének hormontartalmú váladékából, akkor anyává fejlődik, ha nem kap ebből eleget, dolgozóvá. Csak az anyák termékenyek, ha egy család elveszíti az anyát, akkor a dolgozók egyike esetleg elkezdhet megtermékenyítetlen petéket rakni (ő ugye sohasem párzott), de attól a család kihal, így ez általában zsákutca, a család génjei élhetnek esetleg tovább, ha a lerakott petékből kikelő herék párosodnak egy anyával, de nyilván egy méhészen ez nem segít.
Az állatvilágban több a miénktől eltérő rendszer létezik a nemmeghatározásra, ezek egyike a mézelő méh által alkalmazott. Az anya életében egyszer párzik, a begyűjtött spermiumokat egész életében a testében tárolja, amikor petét rak, eldöntheti, hogy megtermékenyíti, vagy nem. A megtermékenyítetlen petékből haploid herék (hímek) kelnek ki, a megtermékenyített petékből lesznek a diploid anyák és dolgozók. Őköztük annyi a különbség, hogyha egy lárva a fejlődése során nagy mennyiségben kap a méhek garatmirigyének hormontartalmú váladékából, akkor anyává fejlődik, ha nem kap ebből eleget, dolgozóvá. Csak az anyák termékenyek, ha egy család elveszíti az anyát, akkor a dolgozók egyike esetleg elkezdhet megtermékenyítetlen petéket rakni (ő ugye sohasem párzott), de attól a család kihal, így ez általában zsákutca, a család génjei élhetnek esetleg tovább, ha a lerakott petékből kikelő herék párosodnak egy anyával, de nyilván egy méhészen ez nem segít.
Azonban létezik egy Apis mellifera capensis nevű jószág, ez a mézelő méh egy alfaja, amely nem meglepő módon Afrikában él, Capetown környékén. A nemmeghatározás itt is ugyanolyan, mint a többi méhnél, viszont ha egy család elveszíti az anyát, akkor egyes dolgozók elkezdenek petéket rakni, amelyekben szűznemzéssel létrejött diploid dolgozók bújnak ki. Ez a szűznemzés úgy történik, hogy a szokásos módon zajló meiózis után a keletkező négy leánysejtmagból kettő összeolvad. Időnként megjelennek aszexuálisan szaporodó A.m. capensis populációk a szexuálisan szaporodó családokban, de ezek általában kis hányadát teszik ki a családnak. Ezek a szaporodóképes A.m. capensis egyedek állandóan nehézségeket okoznak az afrikai méhészeknek. Ugyanis ezek megfertőzik egy másik alfaj, az A.m. scutellata családjait. Ezek a fertőzésnek jellegzetes tüneteket mutatnak: Csökken a hordás. A kaptár bejáratánál scutellata és capensis dolgozók harcolnak. Ahogy elszaporodnak a családban az A. m. capensis egyedek, a család egyre nehezebben boldogul, mert a capensis egyedek valamiért nem gondozzák a fiasítást. A fertőzött család egyre fogyatkozik, míg csak pár capensis lézeng benne, aztán azok is kihalnak. Mivel a dolgozók bejuthatnak más kaptárakba és megtelepedhetnek ott, a fertőzés hamar átterjed a környező családokra is. Különösen rossz a helyzet a méhészetekben, ahol rengeteg család él egymás közvetlen közelében, ráadásul a méhészek gyakran cserélgetnek méheket az egyes családok között. Meglepő módon az A. m. capensis családok kétszer gyakrabban rajzanak, mint az A.m. scutellata családok, ezek a rajok gyakran beolvadnak más családokba, ezzel is terjesztve a fertőzést. A fertőzés vége felé a valódi anya általában eltűnik, de ebben a közleményben még ennek a magyarázatát nem ismerték. (Neumann és Moritz 2002)
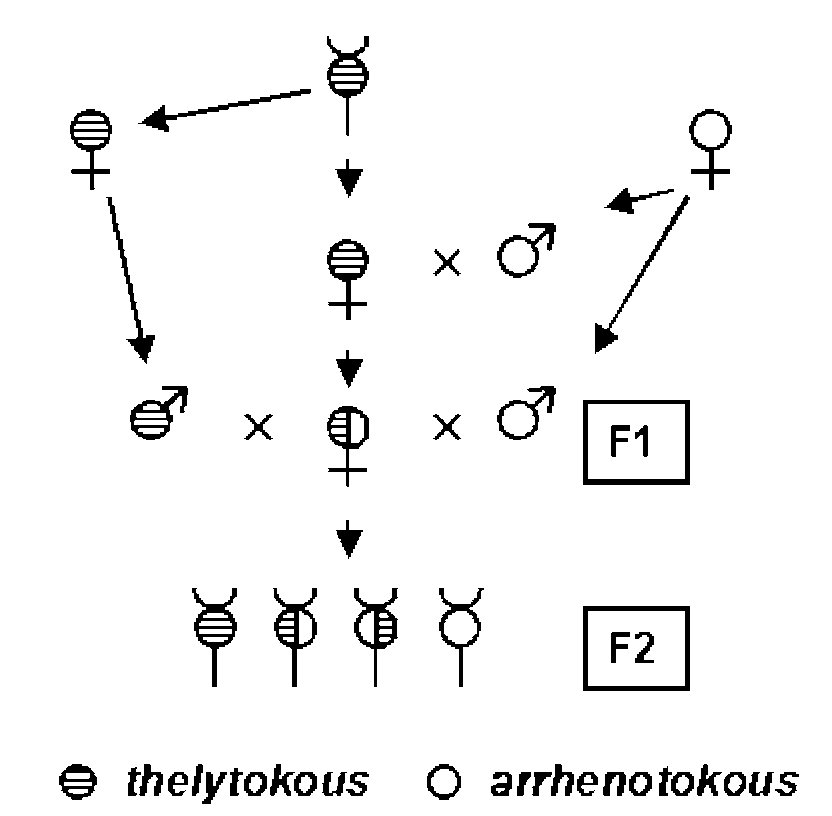 Lattorf és munkatársai 2005 -ben a szűznemzés genetikai hátterét vizsgálták. Hogy eldönthessék, hány gén felelős ennek a jellegnek a kialakításáért, keresztezéseket végeztek. Afrikából A. m. capensis anyákat szereztek, amelyeket először A. m. carnica hímekkel pároztattak (P nemzedék). Az utódaikat (F1) pároztatták A. m. capensis és A. m. carnica hímek 1:1 arányban kevert spermájával termékenyítették meg. Ezek az anyák nyilván heterotigóták bármely génre nézve, amely ennek a jellegnek a kialakításához szükséges így az ő utódaikat (F2) vizsgálták meg. Ez a legegyszerűbb Mendel által is használt tesztkeresztezés, ami itt azért egyszerűbb, mert a hímek haploidok, tehát nem lehetnek heterozigóták. Ezt mutatja az első ábra. Az utódokat egyesével külön ketrecbe tették, megjelölték és kaptak néhány A. m. carnica dolgozót, majd figyelték, hogy petéznek -e?
Lattorf és munkatársai 2005 -ben a szűznemzés genetikai hátterét vizsgálták. Hogy eldönthessék, hány gén felelős ennek a jellegnek a kialakításáért, keresztezéseket végeztek. Afrikából A. m. capensis anyákat szereztek, amelyeket először A. m. carnica hímekkel pároztattak (P nemzedék). Az utódaikat (F1) pároztatták A. m. capensis és A. m. carnica hímek 1:1 arányban kevert spermájával termékenyítették meg. Ezek az anyák nyilván heterotigóták bármely génre nézve, amely ennek a jellegnek a kialakításához szükséges így az ő utódaikat (F2) vizsgálták meg. Ez a legegyszerűbb Mendel által is használt tesztkeresztezés, ami itt azért egyszerűbb, mert a hímek haploidok, tehát nem lehetnek heterozigóták. Ezt mutatja az első ábra. Az utódokat egyesével külön ketrecbe tették, megjelölték és kaptak néhány A. m. carnica dolgozót, majd figyelték, hogy petéznek -e? 
Az utódokat DNS markerek alapján is azonosítani tudták, így meg tudták mondani, melyik egyed melyik apától származott. Az eredmény a második ábrán látható. A két sor az egyik illetve a másik apától származó utódokat jelenti, az oszlopok pedig az összes utódot mutatják (Total), azt, hogy ezekből hányan nem raktak petét egyáltalán (Nonlaying), hányan raktak petét (Laying), és ezek közül hányan raktak diploid petéket, amikből dolgozók keltek ki (Thelytokous) és hányan raktak petéket, amikből csak hímek keltek ki (Arrhenotokous). Ami ebből a kísérletből egyértelműen látszik, hogy az A. m. carnica apától származó utódok nem raktak diploid petéket. Ezzel egyértelműen kizárható, hogy a jelleg anyai öröklődést mutatna, hiszen akkor egy ilyen anya minden lányának ugyanígy diploid petéket kellene raknia. Ugyanígy kizárható, hogy a jelleget egyetlen domináns allél okozza, hiszen akkor a heterozigóta nőstények utódainak a fele kellene, hogy mutassa a fenotípust. Az első sort is megnézve, még több adathoz jutunk. A heterozigóta anyák haploid A. m. capensis apáktól származó utódainak kb. a fele petézett, ezeknek egy része haploid, másik része diploid petéket rakott. Érdekes módon átmenetet nem figyeltek meg, egy egyed vagy csak haploid, vagy csak diploid utódokat hozott létre. Viszont ha a fenotípus kialakításáért egyetlen gén recesszív allélja felelős, akkor ebben a keresztezésben 1:1 hasadási arányt kellene látni, hiszen egy heterozigóta (Aa) nőstény egy A. m. capensis hímtől (a-) származó utódainak a fele heterozigóta (Aa), a fele homozigóta recesszív (aa). A mért eredmények elég jól illeszthetőek erre a modellere, azok az egyedek, amelyek nem petéznek, nem tudni, milyen genotípusúak, de a petéző egyedek kb. fele diploid, másik fele haploid petéket rak, tehát elég valószínű, hogy egyetlen gén recesszív allélja hozza létre ezt a fenotípust. Ha pedig egynél több gén okozná ezt a jelleget, más hasadási arányokat kellene látni. Természetesen nem lehet kizárni, hogy valójában valamilyen sokkal bonyolultabb, sokgénes öröklésről van szó ez esetben, de erre utaló adatot eddig nem találtam.
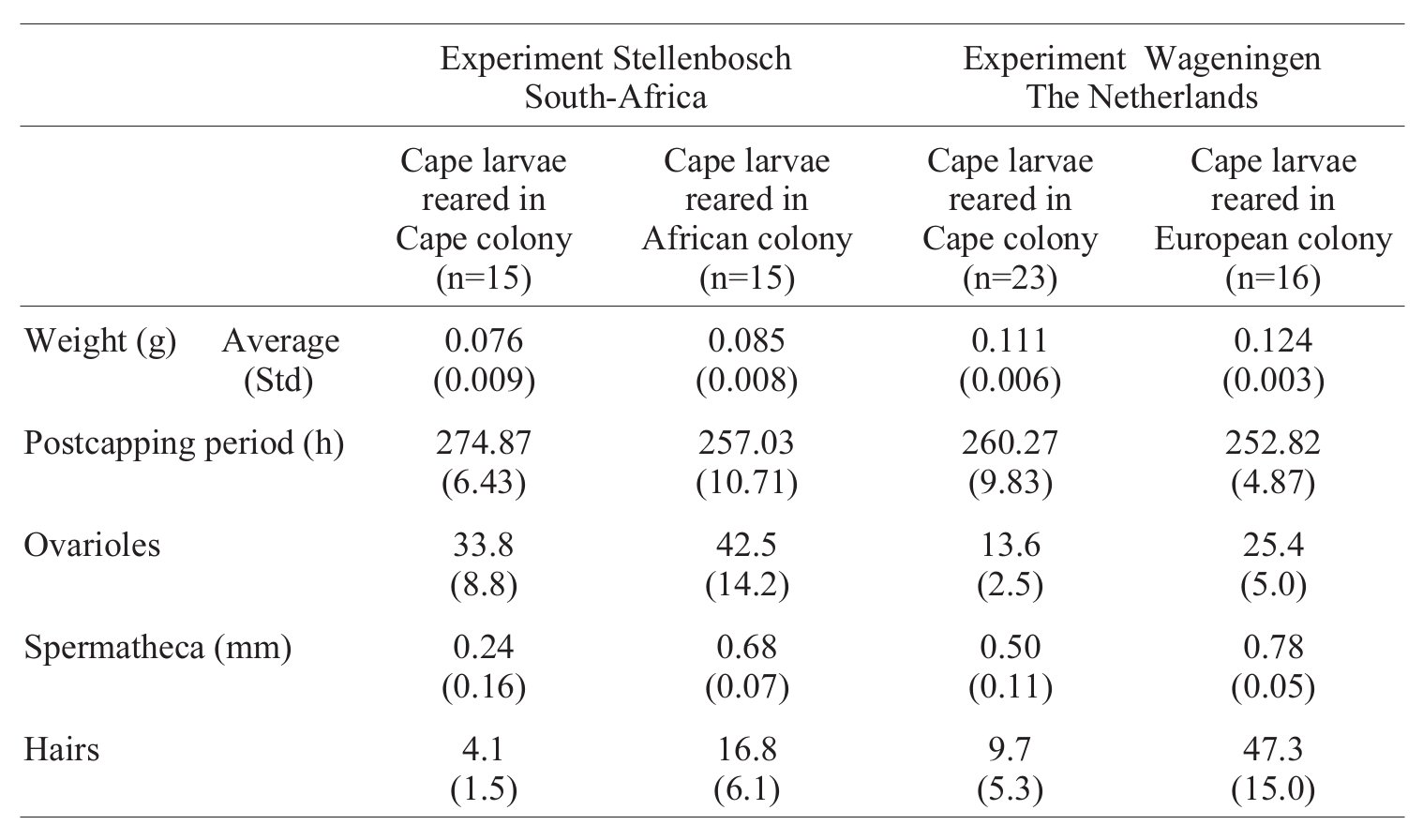
Calis és munkatársai 2001 -ben azt vizsgálták, a táplálék minősége milyen hatással van az A. m. capensis lárvákra. Azt már az elején említettem, hogy az anya és a dolgozók is megtermékenyített petékből kelnek ki, diploidok, a különbség a táplálékukban van, a magas hormontartalmú, sok cukrot tartalmazó méhpempővel etetett lárvákból anya lesz, a kevesebb cukrot és hormont evő lárvákból dolgozó. Ezt mesterségesen is elő lehet idézni, ha juvenilis hormonnal vagy cukrokkal egészítették ki a lárvák táplálékát, akkor mindenféle átmeneti alakok bújtak ki a bábból, vagyis dolgozók, amelyek az anyák egyes jellegzetességeit is mutatták. Megfigyelték, hogy az A. m. capensis lárvák másik alfaj fiasításában több ételt kapnak, mint azok lárvái, így ezekből gyakrabban fejlődnek mindenféle dolgozó-anya köztes alakok. Ezt mutatja a harmadik ábra, A. m. capensis lárvák fejlődését követték nyomon a saját alfajuk kaptáraiban és két helyen két különböző alfaj kaptáraiban, Afrikában és Hollandiában, mindkét kísérletben ugyanazt tapasztalták: Ha az A. m. capensis lárvákat a saját alfajukba tartozó dolgozók gondozzák, szokásos méretűek lesznek, de másik alfaj kaptáraiban gyorsabban fejlődnek, nagyobbra nőnek, nagyobbak lesznek a petefészkeik, és a magtarisznyájuk. Ez lehet a magyarázata, hogy ezek a szocioparaziták miért nem gondozzák a fiasítást, egyszerűen anyaként viselkednek. Ugyanez magyarázza, hogy az A. m. capensis családokat miért nem teszik tönkre ezek az élősködő egyedek, ott ugyanis nagy részük dolgozóként viselkedik.
Martin és munkatársai 2002 -ben azt vizsgálták, hogy mi történik a szocioparaziták által lerakott petékkel? A méhek általában elpusztítják a dolgozók által rakott petéket, ha az anya is petézik. Meglepő módon azt tapasztalták, hogy míg az A. m. scutellata dolgozók által lerakott petéket a többi dolgozó hatékonyan távolította el, az A. m. capensis dolgozók által rakott peték jelentős részét nem bántották. Ezt mutatja a következő ábra, a vízszintes tengely az időt mutatja, a függőleges tengely az, hogy a lerakott peték hány százaléka található még meg. 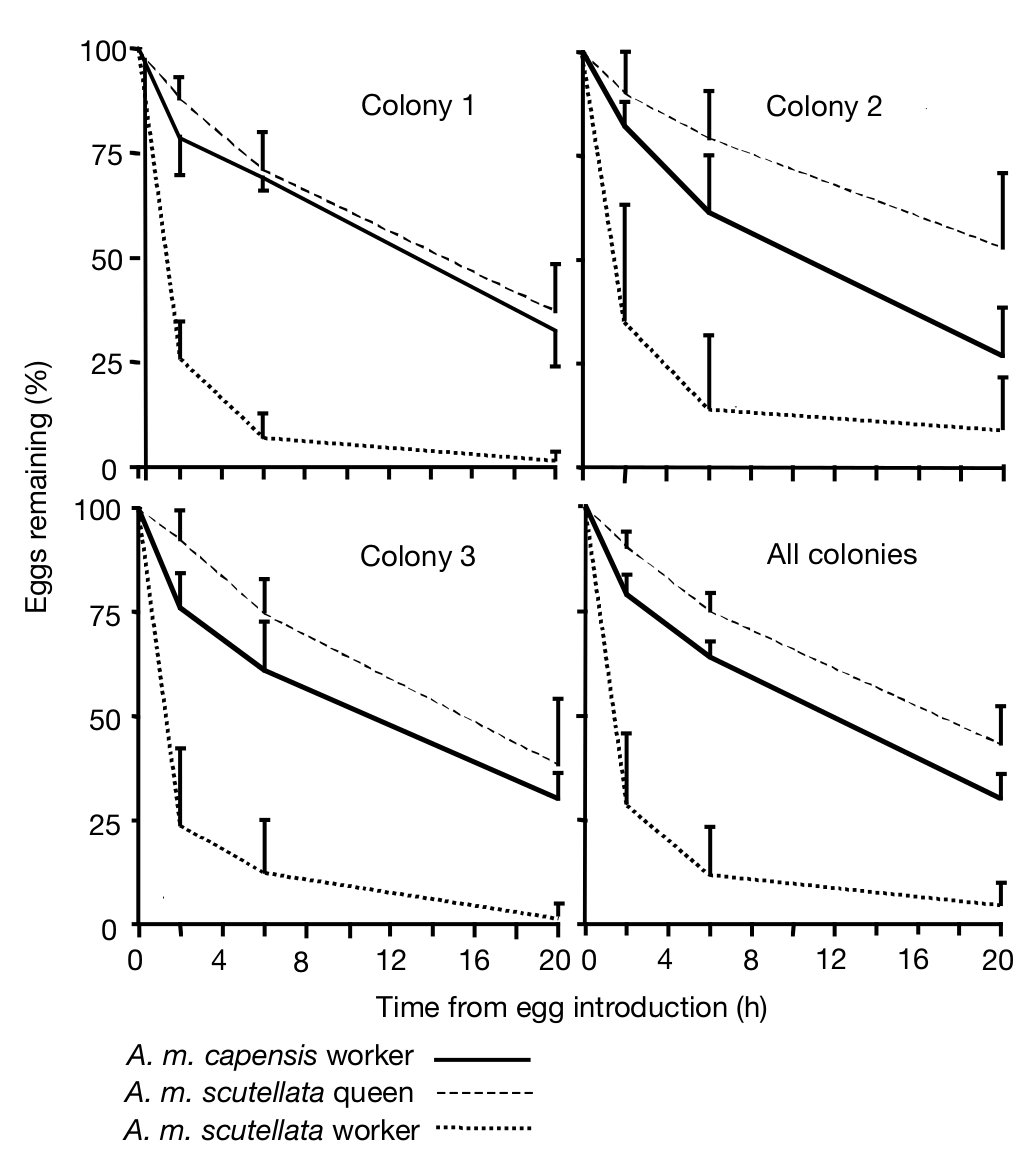
Mennyire veszélyes ez az A. m. capensis élősködő? Eddig csak Afrika déli részein fordult elő, de ahogy behurcolták a scutellata alfaj elterjedési területére, ugyanúgy nyilván behurcolható bárhová. Az európai méhek között éppen úgy képes lenne pusztítani ez a jelenség, de eddig nem fordult elő errefelé, egyelőre csak az afrikai méhészek gondja, de mindenképpen egy viszonylag friss méhkártevőről van szó, amire érdemes odafigyelni a mellett is, hogy biológus szemmel nézve hihetetlenül érdekes maga a jelenség.
Neumann P, Moritz RFA (2002):The Cape honeybee phenomenon: the sympatric evolution of a social parasite in real time? Behav Ecol Sociobiol 52:271–281
Lattorff HM, Moritz RF, Fuchs S. (2005): A single locus determines thelytokous parthenogenesis of laying honeybee workers (Apis mellifera capensis). Heredity 1–5
Calis JNM, Boot WJ, Allsopp MH, Beekman M (2002): Getting more than a fair share: nutrition of worker larvae related to social parasitism in the Cape honey bee Apis mellifera capensis. Apidologie 33 193–202
Martin SJ, Beekman M, Wossler TC, Ratnieks FL. (2002): Parasitic Cape honeybee workers, Apis mellifera capensis, evade policing. Nature.415(6868):163-5.