 Mutációkról már volt szó korábban is, a mai cikkben azonban azt vizsgálták, hogy hogyan viszonyulnak egymáshoz a különböző típusú mutációk. Arról már számos adat ismert, hogy ha egy erős a kompetíció, akkor azok a baktériumsejtek kerülnek előnybe, amelyekben több mutáció történik, azonban a ma boncolgatott cikkben Fehér Tamás és munkatársai azt vizsgálták meg, hogyan kompetál egymással két baktériumtörzs, amelyekben két különböző típusú mutáció történik nagy gyakorisággal.
Mutációkról már volt szó korábban is, a mai cikkben azonban azt vizsgálták, hogy hogyan viszonyulnak egymáshoz a különböző típusú mutációk. Arról már számos adat ismert, hogy ha egy erős a kompetíció, akkor azok a baktériumsejtek kerülnek előnybe, amelyekben több mutáció történik, azonban a ma boncolgatott cikkben Fehér Tamás és munkatársai azt vizsgálták meg, hogyan kompetál egymással két baktériumtörzs, amelyekben két különböző típusú mutáció történik nagy gyakorisággal.
Mutációkról már volt szó korábban is, a mai cikkben azonban azt vizsgálták, hogy hogyan viszonyulnak egymáshoz a különböző típusú mutációk. Arról már számos adat ismert, hogy ha egy erős a kompetíció, akkor azok a baktériumsejtek kerülnek előnybe, amelyekben több mutáció történik, azonban a ma boncolgatott cikkben Fehér Tamás és munkatársai azt vizsgálták meg, hogyan kompetál egymással két baktériumtörzs, amelyekben két különböző típusú mutáció történik nagy gyakorisággal.
Hogy ezt a kérdést egyáltalán megközelíthessék, felhasználták a Pósfai György laborjában előállított MDS42 Escherichia coli baktériumtörzset, amelynek a genomjából eltávolítottak számos nem-létfontosságú gént. Ez számokban kifejezve annyit jelent, hogy a vad típusú E. coli törzs 4434 génjéből eltávolítottak 743 -t, ami a genom valamivel több, mint tizenöt százalékát jelenti. Ezek mellett eltávolították a genomból az összes ugráló genetikai elemet is, így ezek mutációkat sem okoznak ebben a törzsben.
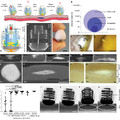
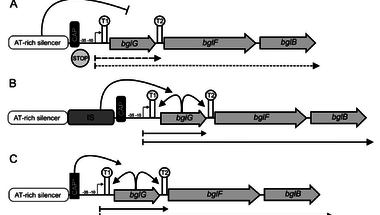
A kísérletek során a baktériumokat minimál táptalajon tartották, egyetlen szénforrásként szalicint adva hozzá. Az E. coli ugyanis alapesetben képtelen felhasználni a szalicint, bár megvannak a genomjában az ehhez szükséges enzimeket kódoló gének, ám ez a bgl operon nem fejeződik ki és nem is indukálható. Így a szalicintartalmú minimáltáptalajra szélesztett E. coli sejtek túlnyomó többsége éhen hal, egyedül a ritka, újonnan megjelenő, szalicint felhasználni képes mutánsok maradnak meg. Ezek láthatóak az első ábrán. Alapesetben a bgl operon kifejeződését egy AT gazdag represszorszekvencia gátolja és ha mégis elindulna a transzkripció, a T1 és a T2 terminátor szakaszok le is állítják. Ha egy ugráló genetikai elem épül be a represszorszekvencia mellé (B ábra), a bglG gén átíródik, az általa kódolt fehérje szalicin jelenlétében képes meggátolni a két terminátorszakasz működését, így átíródik az operon, a sejt fel tudja használni a szalicint. E mellett számos pontmutáció képes aktiválni a bgl operont, hiszen mutációk nyomán működésképtelenné válhat a represszorszekvencia és a két transzkripciós terminátorszekvencia is. Ebben a rendszerben a bgl operont aktiváló mutációk egyértelműen előnyösek, amelyik baktériumsejtben megtörténnek, az nem éhezik tovább, így könnyen túlnövi azokat, amelyekben nem képesek felhasználni a szalicint.
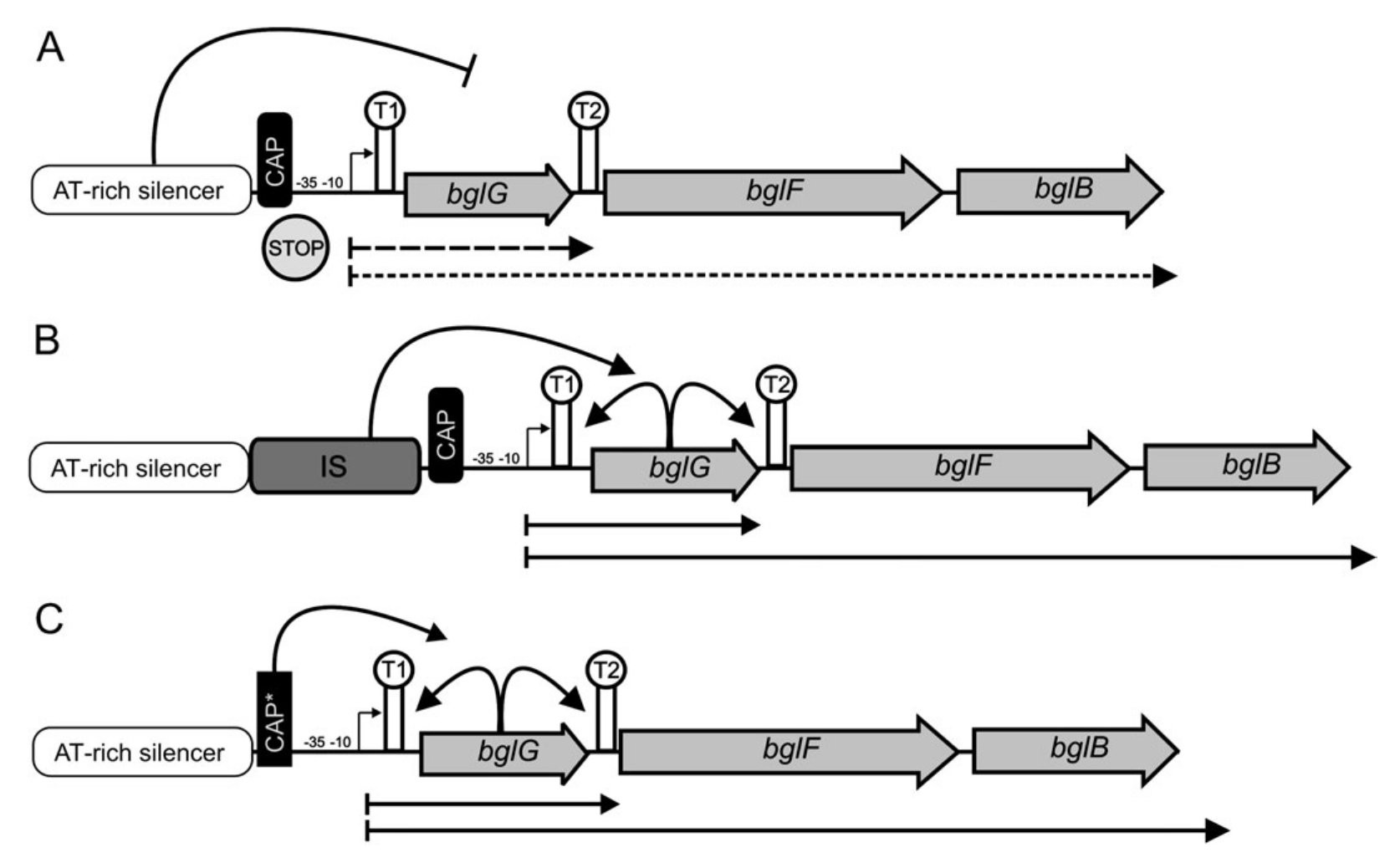
 A kísérlet másik eleme az egyes baktériumtörzsek felismerése, hiszen ahhoz, hogy tudjuk, melyik törzs nőtte túl a másikat, valahogyan el kell különíteni egymástól a két törzset. Az egyes törzsek genomjába éppen ezért bejuttattak egy kéken fluoreszkáló fehérjét (CFP) kódoló gént vagy egy sárgán fluoreszkáló fehérjét (YFP) kódoló gént, így egyszerűen ibolyántúli fényben megkülönböztethető egymástól a kéken és a sárgán világító két törzs. Így egy kevert kultúrában megállapítható az egyik illetve a másik törzsből származó sejtek pontos aránya. Ez látható a második ábrán. Ráadásul a sejtek nem folyamatosan fejezték ki ezeket a géneket, a leolvasás előtt anhidotetraciklinnel indukálták a CFP és YFP fehérjék kifejeződését, így azok nem zavarhatták meg a sejtek növekedését. Így összesen négy baktériumtörzzsel dolgoztak az MDS42 törzshöz viszonyították a többit, az MDS40-IS1 -t, amely egyetlen példányt tartalmaz az IS1 ugráló genetikai elemből, az MDS42-ΔmutS -t, amely egy mutátor törzs, sokkal gyakrabban történnek benne pontmutációk, mint az MDS42 -ben és az MDS42-IS1-ΔmutS -t, amelyben a pontmutációk is gyakoribbak és egy ugráló genetikai elemet is tartalmaz.
A kísérlet másik eleme az egyes baktériumtörzsek felismerése, hiszen ahhoz, hogy tudjuk, melyik törzs nőtte túl a másikat, valahogyan el kell különíteni egymástól a két törzset. Az egyes törzsek genomjába éppen ezért bejuttattak egy kéken fluoreszkáló fehérjét (CFP) kódoló gént vagy egy sárgán fluoreszkáló fehérjét (YFP) kódoló gént, így egyszerűen ibolyántúli fényben megkülönböztethető egymástól a kéken és a sárgán világító két törzs. Így egy kevert kultúrában megállapítható az egyik illetve a másik törzsből származó sejtek pontos aránya. Ez látható a második ábrán. Ráadásul a sejtek nem folyamatosan fejezték ki ezeket a géneket, a leolvasás előtt anhidotetraciklinnel indukálták a CFP és YFP fehérjék kifejeződését, így azok nem zavarhatták meg a sejtek növekedését. Így összesen négy baktériumtörzzsel dolgoztak az MDS42 törzshöz viszonyították a többit, az MDS40-IS1 -t, amely egyetlen példányt tartalmaz az IS1 ugráló genetikai elemből, az MDS42-ΔmutS -t, amely egy mutátor törzs, sokkal gyakrabban történnek benne pontmutációk, mint az MDS42 -ben és az MDS42-IS1-ΔmutS -t, amelyben a pontmutációk is gyakoribbak és egy ugráló genetikai elemet is tartalmaz.
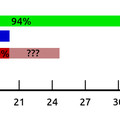
Az első kísérletben MDS42 és MDS42-IS1 törzseket kevertek össze egyenlő arányban, majd egy hétig hagyták őket evolválni, aztán megmérték, hogy a szalicint emészteni képes sejtek milyen arányban származnak az egyik illetve a másik törzsből. Azt tapasztalták, hogy kilencszer több szalicint emészteni képes egyed származik az MDS42-IS1 törzsből, az egyetlen ugráló genetikai elem jelenlétét a bgl operonban a szalicinemésztő törzsek kilenctizedéből ki is tudták mutatni. Ezek után ugyanebben a rendszerben megvizsgálták az MDS42-ΔmutS törzset, amelyben az MDS42 -höz képest százszor több pontmutáció történik. Azt tapasztalták, hogy ilyen körülmények között az egy hétig evolválni hagyott sejtek között minden hatszáznyolcvanhatodik mutátor törzsból származóra jutott csak egy MDS42 törzsből származó, megállapították, hogy a mutátor törzs elég nagy előnyt élvez ezekben a kísérletekben. Ezek után megpróbálták az MDS42-IS1 törzset a MDS42-ΔmutS törzzsel evolváltatni és azt tapasztalták, hogy a kísérlet végére a szalicinemésztő sejtek 1:2,4 arányban származtak az MDS42-IS1 és a MDS42-ΔmutS törzsből, ebből úgy tűnik, hogy a pontmutációk előnyösebbek, mint az ugráló genetikai elemek beépülései. Ezek után kézenfekvő a kérdés, hogy ha az ugráló genetikai elem jelenléte is előnyös és a mutátor fenotípus is előnyös, mi történik, ha egy sejt mutátor fenotípust is mutat és egy ugráló genetikai elemet is tartalmaz? Ezt a kísérletet is elvégezték, az MDS42-IS1-ΔmutS törzs sikeresen kompetál az MDS42-IS1 és a MDS42-ΔmutS törzzsel is. Ezen eredmények láthatóak a harmadik ábrán.
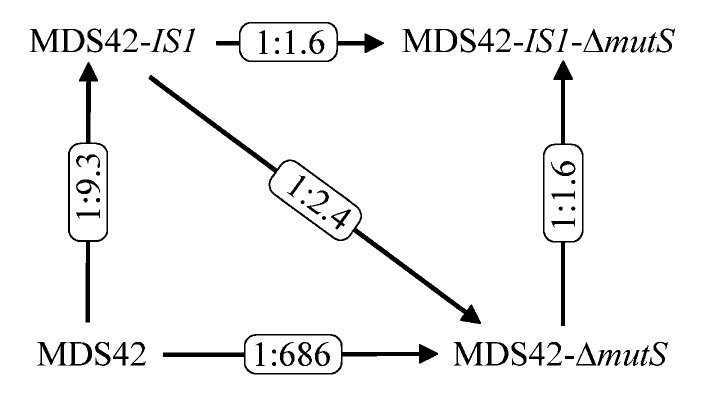
No most, milyen következtetéseket lehet levonni ezekből a kísérletekből? Ha egy élőlénynek új környezethez kell alkalmazkodnia, a magasabb mutációs ráta egyértelműen előnyös, de úgy tűnik nem mindegy, milyen mutációk történnek: Ebben a rendszerben a pontmutációk egyértelműen előnyösebbek mint egy ugráló genetikai elem mozgása.
Fehér T, Bogos B, Méhi O, Fekete G, Csörgo B, Kovács K, Pósfai G, Papp B, Hurst LD, Pál C. (2012): Competition between Transposable Elements and Mutator Genes in Bacteria. Molecular Biology and Evolution [Epub ahead of print]
Pósfai G, Plunkett G 3rd, Fehér T, Frisch D, Keil GM, Umenhoffer K, Kolisnychenko V, Stahl B, Sharma SS, de Arruda M, Burland V, Harcum SW, Blattner FR. (2006): Emergent properties of reduced-genome Escherichia coli. Science. 312(5776):1044-6.