Ma nyomon követjük, hogyan is terjednek el az evolúciótagadás népmesei elemei a világháló bugyraiban, egyszersmind megláthatjuk, hogyan is születnek az „evolúciót cáfoló” bizonyítékok.
A kérdéses közlemény címe: Hotspots of Biased Nucleotide Substitutions in Human Genes. Hagyományosan abból szokás kiindulni, hogy ha két, rokon élőlény genomjában olyan helyeket találnak, amelyek a genom átlagos mutációs gyakoriságánál több mutációt szedtek össze, akkor erre a génre pozitív szelekciós nyomás hatott, hiszen gyorsabban változott, mint ahogy a genetikai sodródás indokolta volna. Vannak matematikai módszerek, amivel azonosíthatóak az ilyen szakaszok a genomon belül.
Ma nyomon követjük, hogyan is terjednek el az evolúciótagadás népmesei elemei a világháló bugyraiban, egyszersmind megláthatjuk, hogyan is születnek az „evolúciót cáfoló” bizonyítékok.
A kérdéses közlemény címe: Hotspots of Biased Nucleotide Substitutions in Human Genes. Hagyományosan abból szokás kiindulni, hogy ha két, rokon élőlény genomjában olyan helyeket találnak, amelyek a genom átlagos mutációs gyakoriságánál több mutációt szedtek össze, akkor erre a génre pozitív szelekciós nyomás hatott, hiszen gyorsabban változott, mint ahogy a genetikai sodródás indokolta volna. Vannak matematikai módszerek, amivel azonosíthatóak az ilyen szakaszok a genomon belül.
Az régi megfigyelés, hogy vannak a genomban olyan pontok, ahol gyakoriak a rekombinációs események, ráadásul ezeken a területeken gyakoribbak a G és C bázispárok, mint általában a genom egészében, feltételezik, hogy a gyakori átkereszteződések felelősek ennek az egyenlőtlenségnek a kialakításáért. Jó pár bizonyíték szól e magyarázat mellett:
-Ha egymáshoz illesztjük az emberi és majom genomokat, a gyakori A,T -> G,C báziscsere típusú különbségek egybeesnek az emberben gyakran rekombinálódó genomi szakaszokkal.
-Az emlős és madár genomok gyakran rekombinálódó szakaszai, például az X kromoszómák pszeudoautoszómás régiói, vagy a duplikálódott szakaszok mind különösen GC gazdagok.
-A genom GC tartalma számos eukarióta szervezetben korrelál a rekombináció gyakoriságával.
-Sejtvonalakon végzett kísérletek azt mutatják, hogy a DNS repair mechanizmusok nagyobb gyakorisággal építenek be G vagy C nukleotidokat egy rosszul párosodó bázispár helyére, mint A -t vagy T-t.
Ezek a területek jól megkülönböztethetőek a genom többi részétől, ahol gyakoribbak a G,C -> A,T típusú báziscsere mutációk. Ezek a bizonyos rekombinációs hotspotok viszonylag kis méretű genomi szakaszok, jellemzően az egy kilobázisnyi méretet sem érik el és nagyon rövid életűek. Éppen ezért feltételezik, hogy a gyakori rekombinációs események nyomán működő DNS repair mechanizmusok működése okozza ezeken a területeken a G és C nukleotidok felszaporodását.
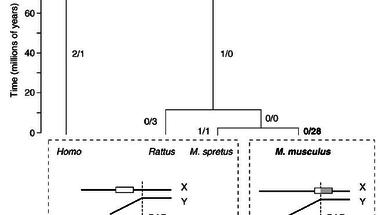
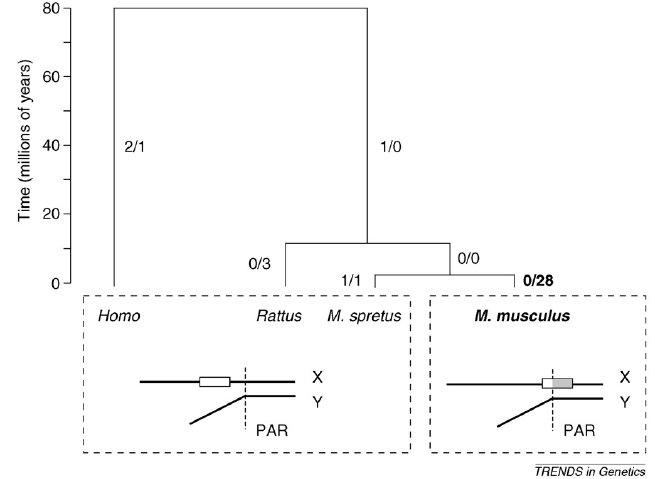
Egy példát is hoznak erre, az Fxy gén a Mus spretus nevű egérben, a patkányban és az emberben is az X kromoszómán található, ám a Mus musculus nevű egérben ez a gén részben átkerült az X-kromoszóma pszeudoautoszómás régiójába, ahol gyakoriak a rekombinációs események. A gén 5' szakasza egyetlen bázispárban tér el a Mus musculus és a Mus spretus egerekben. Ha összehasonlítjuk a patkány Fxy génjének azonos szakaszával, látszik, hogy a Mus musculus gén szekvenciája egyezik vele, míg a Mus spretusé különbözik tőle egyetlen bázisban. Azonban a gén 3' szakasza, amely a gyakran átkereszteződő régióba került át, a Mus spretusban egyezik a patkány szekvenciával, míg huszonnyolc bázispárban tér el a Mus musculus esetében és minden egyes pontmutáció A, T -> G, C típusú. Ismereteink szerint semmilyen előnyt nem jelentenek ezek a mutációk, e génszakasz gyors változását egyedül az okozza, hogy a gyakori rekombináció miatt itt gyakoriak a báziscsere típusú mutációk. Mi a következménye ennek a rengeteg mutációnak? Az Mus spretus egér Fxy fehérjéje összesen hat aminosavban különbözik az emberétől, a két faj elválása óta (~80 millió éve) ennyi mutációt gyűjtött be. Azonban a M. musculus Fxy fehérje huszonnyolc aminosavban különbözik a M. spretus fehérjétől, pedig ez a két faj csak körülbelül 1-3 millió éve vált szét. Itt bizony több mint százszor gyorsabban rögzülnek mutációk, mint a BGC által nem érintett genomi szakaszokon. Ez látható az ábrán is.
Ebben a közleményben mindössze annyit írnak le, hogy elemezték az egész emberi genomot és megkeresték azokat a szakaszokat, amelyek érint ez a jelenség, vagyis ahol a rekombináció nyomán fölszaporodnak a G, C nukleotidok. Mivel ezek a szakaszok egy kilobázisnál is rövidebbek, legtöbbször egy génnek csak egyetlen exonját érintik. Összesen nyolcvanhárom exon érintett, nyolcvankét emberi génből, ezen exonok átlaghosszúsága 516 bázispár. Ezek a szakaszok átlagosan 7,73 báziscserét tartalmaznak, azaz a szakaszok másfél százalékát jelentik. A báziscserék során 326 A,T -> G,C és 248 G,C -> A,T változás történt, ezek aránya 0,57, míg az egész genomban ugyanez az arány 0,39.
Nem meglepő módon ez a jelenség nem korlátozódik az exonokra, az őket körülvevő intronokban is jellemző, ám minél távolabbi szakaszt vizsgáltak, annál ritkábban történtek ilyen mutációk.
Vannak a genomnak olyan szakaszai, ahol jóval gyakoribbak az A,T -> G,C mutációk, mint a G,C -> A,T báziscserék. Ez szerintem nem különösebben forradalmi eredmény, a mondás mindössze annyi, hogy ha valaki gyorsan változó gént vizsgál, érdemes megnéznie előtte, hogy nem egy ilyen mutációs forró pont hatását látja -e? No most miért is érdekes számunkra ez a cikk? A magyar világhálón önálló életet kezdett élni, az evolúciótagadók egyik érvévé vált.
Az egész egy különösen szenzációhajhász sajtóközleménnyel indult, ami a szerény „Nem csak a természetes kiválasztódás hajtja az evolúciót” címmel íródott. Ez körülbelül illeszkedik a saját kutatóik eredményeit pofátlanul felfújó sajtóközlemények sorába, az egészet úgy állítja be, mintha valamilyen őrült gyakori folyamatot fedeztek volna fel, amely egyedül lenne felelős az ember és a csimpánz közti összes különbségért, holott ez a jelenség összesen nyolcvankét gént érint a huszonháromezerből (0,35% ). Ennek egy magyar fordítása is napvilágot látott.
Ennek az átirata kikerült egy kreacionista oldalra is, ahol valamilyen megfontolásból hozzácsaptak egy teljesen más kérdésről készült interjút is. Itt már át is nemesedett a közlemény az evolúció teljes és végleges cáfolatává, sőt, a hozzászólások között a bloggazda már az összeesküvést is hozzátette, egyértelműen állítja, hogy a szerzők eredetileg azt akarták leközölni, hogy megcáfolták az evolúciót, de nem merték, mert féltették az állásukat. Sőt, még engem is megvádolt, hogy én készakarva elhallgatom az evolúciót tagadó megdönthetetlen bizonyítékokat. Innentől pedig a magyar népmesék közé máris leszűrődött, hogy ez a közlemény bizony az evolúció egyértelmű cáfolata, bár azt tulajdonképpen senki sem tudja megmondani miért is az?
Valójában mi is ennek a jelenségnek a valós súlya? Mekkora részt vállal az evolúcióból a BGC? Vélhetőleg a semminél alig valamivel többet. Egyrészt a genom nagyon rövid szakaszaira jellemző, mint ahogy láttuk az emberi gének töredékszázalékát érinti, azok esetében is csak egy-egy exont.
Milyen „irányba” hajtja az élőlényeket a BGC? Mert addig oké, hogy a G és C nukleotidok szaporodnak föl egy rövid genomi szakaszon, de ez milyen irányba változtatja meg az élőlényeket? Nyilván, mint minden mutációnak, a BGC következtében felszaporodó báziscseréknek sincs egységes hatása, az egyes mutációk fenotípusra gyakorolt hatását nem lehet visszakövetkeztetni ennyi adatból, így akármilyen fenotípusos változásokat okoznak is, ezeknek a változásoknak pont úgy nincs „irányuk”, mint ahogy az összes többi mutációnak sem. Lehetnek előnyösek, hátrányosak, vagy éppen semlegesek az adott körülmények között.
Bármilyen módon akadályozza -e a természetes szelekciót a BGC? Nyilván a két jelenségnek semmi köze sincs egymáshoz, ha egy GC gazdag allél valamiért hátrányos, ugyanúgy szelekciós nyomás hat majd az eltűnésére. Az, hogy valamilyen előnyt élvez egy allél, csak azért, mert több benne a G és a C nukleotid, nyilván csak addig segíti az elterjedését, amíg ezt az előnyt nem múlják felül a hátrányai. Nyilván semleges, vagy nagyon enyhén hátrányos allélek elterjedését segítheti, de ha egy allél jelentősen csökkenti az egyedek fittneszét, akkor hiába GC gazdag, a hordozó egyedek kihalásával el fog tűnni.
Ezek után mi a BGC jelentősége? Tulajdonképpen nem sok, egyszerűen arról van szó, hogy hagyományosan ha két közeli rokon faj genomjában egy adott ponton nagyon sok eltérést tapasztalunk, arra gondolunk, hogy ezek bizony a két faj közti különbségek okai, olyan genomi régiót vizsgálunk, amire nagyon erős szelekciós nyomás nehezedett. A BGC felismerése ezt a képet csak annyiban árnyalja, hogy egy ilyen szakasz vizsgálata előtt érdemes ellenőrizni, hogy nem egy rekombinációs hotspot torzítja -e az eredményeinket? Ez nem cáfolja meg az evolúciót, nem helyezi hatályon kívül a természetes szelekciót. Hogy miért pont ez a közlemény vált ilyen népszerűvé, az csak a rátapadt népmesei elemekkel magyarázható, amiknek viszont semmi alapja sincs.
Galtier N. and Duret L. (2007): Adaptation or biased gene conversion? Extending the null hypothesis of molecular evolution; TRENDS in Genetics Vol.23 No.6
Berglund J., Pollard K. S., Webster M. T. (2008):Hotspots of Biased Nucleotide Substitutions in Human Genes; PLOS biology vol 7. issue 1
Sexcomb