 Pontosabban még nem vizsgát okok miatt (ötletelni persze lehet), a paradigmaváltások szele általában nehezen éri el a tudományos ismeretterjesztést. Mással ugyanis nehezen magyarázható, hogy egy olyan korban, amikor szakmai berkekben a "szemét DNS" ("junk DNA") fogalma, max. tudománytörténeti kuriózumként kerül elő, számos cikk, előadás, stb. még mindig olyan drámaian zengi bele a világba, hogy "a tudósok megfejtették a dzsunkdéjenes titkát", mintha azon komolyan meg kellene lepődjön a publikum.
Pontosabban még nem vizsgát okok miatt (ötletelni persze lehet), a paradigmaváltások szele általában nehezen éri el a tudományos ismeretterjesztést. Mással ugyanis nehezen magyarázható, hogy egy olyan korban, amikor szakmai berkekben a "szemét DNS" ("junk DNA") fogalma, max. tudománytörténeti kuriózumként kerül elő, számos cikk, előadás, stb. még mindig olyan drámaian zengi bele a világba, hogy "a tudósok megfejtették a dzsunkdéjenes titkát", mintha azon komolyan meg kellene lepődjön a publikum.
Pedig a valóság az, hogy minimum 30 éve tudjuk, hogy a genom nem fehérje kódoló részei is fontosak az élőlények fejlődése szempontjából. 1975-ben Marie-Claire King és Allan Wilson vette észre először, hogy az emberi és csimpánz genom 98%-ban azonos (persze nem voltak akkor még teljes genomok, de a rendelkezésükre álló szekvenciák alapján extrapoláltak - mint később kiderült, igencsak pontosan) és ez a kevéske különbség sem elsősorban a gének fehérje kódoló részében rejlik. Ebből született az a felismerés, ami az origóját jelentette a későbbi szemléletváltásnak: "[a két faj] makromolekulái annyira hasonlítanak, hogy szabályozó mutációk lehetnek felelősek biológiai különbségeikért."
Pontosabban még nem vizsgát okok miatt (ötletelni persze lehet), a paradigmaváltások szele általában nehezen éri el a tudományos ismeretterjesztést. Mással ugyanis nehezen magyarázható, hogy egy olyan korban, amikor szakmai berkekben a "szemét DNS" ("junk DNA") fogalma, max. tudománytörténeti kuriózumként kerül elő, számos cikk, előadás, stb. még mindig olyan drámaian zengi bele a világba, hogy "a tudósok megfejtették a dzsunkdéjenes titkát", mintha azon komolyan meg kellene lepődjön a publikum.
Pedig a valóság az, hogy minimum 30 éve tudjuk, hogy a genom nem fehérje kódoló részei is fontosak az élőlények fejlődése szempontjából. 1975-ben Marie-Claire King és Allan Wilson vette észre először, hogy az emberi és csimpánz genom 98%-ban azonos (persze nem voltak akkor még teljes genomok, de a rendelkezésükre álló szekvenciák alapján extrapoláltak - mint később kiderült, igencsak pontosan) és ez a kevéske különbség sem elsősorban a gének fehérje kódoló részében rejlik. Ebből született az a felismerés, ami az origóját jelentette a későbbi szemléletváltásnak: "[a két faj] makromolekulái annyira hasonlítanak, hogy szabályozó mutációk lehetnek felelősek biológiai különbségeikért."
Azóta arra is fény derült, hogy "több biológiai folyamat, mint jelátviteli útvonal", azaz egyes gének termékei a fejlődés és élet során újból és újból felhasználódnak, a legkülönbözőbb szervekben, ami szintén csak a szabályozásuk komplexitásával magyarázható. Így aztán nem csoda, hogy az elmúlt tíz évben rengetgen kezdték az említett szabályozó régiókat vizsgálni, néha egész aprólékosan, hogy megleljék az egyes gének különböző szabályozó elemeit.
Ez a kutakodás aztán magasabb sebességi folyamatba kapcsolt, ahogy egyre több faj genomját megszekvenálták: a teljes genom összehasonlítások ugyanis a napnál is világosabban mutatták, hogy léteznek olyan nemkódoló DNS részek, amelyek alig-alig változtak az év százmilliók során, és szinte bázispárra pontosan úgy néznek ki egy halban, mint bennünk, emberekben. Ezeket a mai, letisztultabb szakirodalom CNE-ként aposztrofálja ("conserved non-coding elements"), bár konvenció kialakulása előtt, kacifántosabb rövidítéseket is lehetett találni, mint pl. MFCS ("mammals-fishes-conserved-sequence").
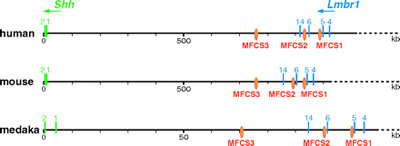 Hogy miként is működnek ezek a CNE-k, arra talán a legjobb példa az egyik, fejlődésbiológiai igen fontos gén, a sonic hedgehog (shh) szabályozórégiója.
Hogy miként is működnek ezek a CNE-k, arra talán a legjobb példa az egyik, fejlődésbiológiai igen fontos gén, a sonic hedgehog (shh) szabályozórégiója.
A shh központi szerepet játszik egyebek mellett a végtag- és szemfejlődésben, a fogak kialakulásában, hogy csak a blog által korábban lefedett területeket említsem. A különböző folyamatokat eltérő szabályozó elemek irányítják, amelyek a géntől több száz kilobázis távolságra helyezkednek el, két CNE (MFCS1 és -2) esetében egy szomszédos gén, az lmbr1 nem fehérjekódoló részeiben, intronjaiban. (Ez nem különlegesség, hanem igen gyakori, jól példázva hogy a.) miért is nem triviális a gén fizikai definíciója, ill. b.) miért fontosak a funkcionális vizsgálatok, hiszen pusztán szekvenciaösszehasonlítás alapján hihetnénk azt is, hogy az említett régiók az lmbr1 szabályozórégiói.)
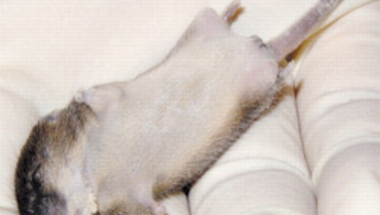
A shh teljes hiánya/működésképtelensége, pont a gén sokoldalúsága miatt, súlyosan halálos. A fejlődő embrió számos szerve rosszul, vagy sehogy sem alakul ki, esély sincs az életbenmaradásra. Azonban, ha csak az MFCS1 szekvenciáját "lopjuk ki" egy egérből, akkor egy életképes egérhez jutunk, ám ennek a végtagjai súlyosan deformáltak lesznek.
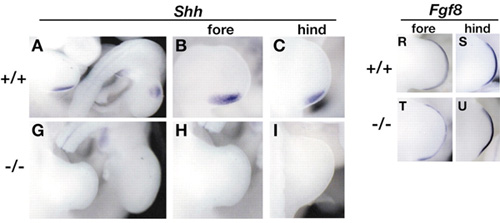 Ha a végtagok fejlődését nézzük meg tüzetesen (és itt most újfent őszintén ajánlanám a korábbi végtagfejlődésről szóló posztot tanulmányozásra), azt láthatjuk, hogy egy MFCS1 mutánsban ("-/-") a végtagbimbó kezdeti fejlődését biztosító fgf gének kifejeződése teljesen normális, azonban sem a mellső, sem a hátsó végtagokban ("fore-" és "hindlimb") nem jelenik meg a végtagfejlődés későbbi fázisait irányító shh.
Ha a végtagok fejlődését nézzük meg tüzetesen (és itt most újfent őszintén ajánlanám a korábbi végtagfejlődésről szóló posztot tanulmányozásra), azt láthatjuk, hogy egy MFCS1 mutánsban ("-/-") a végtagbimbó kezdeti fejlődését biztosító fgf gének kifejeződése teljesen normális, azonban sem a mellső, sem a hátsó végtagokban ("fore-" és "hindlimb") nem jelenik meg a végtagfejlődés későbbi fázisait irányító shh.
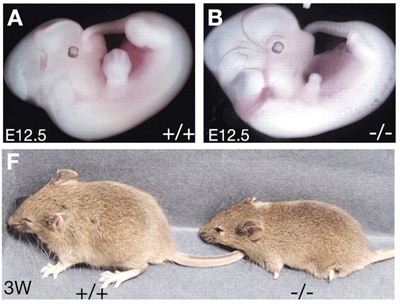 Ennek a következménye aztán az elsatnyult végtag, ami már az embrionális fejlődés közepén is evidens, és mire az egerek világra jönnek teljesen egyértelművé válik.
Ennek a következménye aztán az elsatnyult végtag, ami már az embrionális fejlődés közepén is evidens, és mire az egerek világra jönnek teljesen egyértelművé válik.
Vagyis szabályozószekvenciák kaialkulása vagy megszűnése révén, folyamatspecifikusan ki ill. be kapcsolhatunk géneket, anélkül, hogy más funkcióiknak keresztbetennénk. Ez egy borzasztóan fontos jellegzetesség, ami aztán hatalmas "játszóteret" biztosít az evolúció számára is.
Például, bár a shh MFCS1-e gyakorlatilag minden gerinces fajban fellelhető (egy japán gömbhalban épp úgy, mint egy emberben), pont a végtag nélküli gilisztagőtében ill. számos kígyófajban nincs jelen. Hogy ez a hiány a lábatlanság oka vagy egyszerűen csak okozata, az hosszan vitatható lenne, bár a kígyók esetében minden valószínűség szerint az utóbbi. Az ősi, már csak csökevényes lábakat hordó kígyókban, nem "őrködött" tovább a szelekció a MFCS1 szekvenciája felett és végül az is "elcsökevényesedett", a felismerhetetlenségig mutálódott.
Sagai T, Hosoya M, Mizushina Y, Tamura M, Shiroishi T (2005) Elimination of a long-range cis-regulatory module causes complete loss of limb-specific Shh expression and truncation of the mouse limb. Development 132(4): 797-803.
Sagai T, Masuya H, Tamura M, Shimizu K, Yada Y, et al. (2004) Phylogenetic conservation of a limb-specific, cis-acting regulator of Sonic hedgehog (Shh). Mamm Genome 15(1): 23-34.