(Vendégmunkásunk, SexComb, újra lesújt.)
(Vendégmunkásunk, SexComb, újra lesújt.)
Cikksorozatunk harmadik részéhez érkezett (előzmények itt és itt), szerencsére az értelmes tervezés hívei ellátnak bőven elemeznivalóval. A ma boncasztalra fektetett cikk itt található, a jelöletlen idézetek ebből az írásból származnak.
Mendelt nem véletlenül tartjuk a genetika atyjának. Az őáltala végzett kísérleteket a mai napig tanítják az egyetemeken, éppen azért, mert tiszta kísérleti rendszert hozott létre, adatokat gyűjtött, majd az eredményeket megfelelően értékelte ki. Mit is csinált ez a szerzetes? Először is megfelelő modellszervezetet keresett, amelynek végül is a borsó, azaz a Pisum sativum bizonyult. A borsó ugyanis képes önmegtermékenyítésre, gyorsan nő, kevés helyet igényel, ráadásul, mivel termesztett növény, rengeteg változata ismert. Mendel harminckét borsótörzsből két év alatt választotta ki azt a néhány vonalat, amellyel kísérletezni kezdett. Ezek után hét tulajdonságot választott ki, a további munkája során ezek öröklődését vizsgálta.
(Vendégmunkásunk, SexComb, újra lesújt.)
Cikksorozatunk harmadik részéhez érkezett (előzmények itt és itt), szerencsére az értelmes tervezés hívei ellátnak bőven elemeznivalóval. A ma boncasztalra fektetett cikk itt található, a jelöletlen idézetek ebből az írásból származnak.
Mendelt nem véletlenül tartjuk
a genetika atyjának. Az őáltala végzett kísérleteket a mai napig
tanítják az egyetemeken, éppen azért, mert tiszta kísérleti rendszert
hozott létre, adatokat gyűjtött, majd az eredményeket megfelelően
értékelte ki. Mit is csinált ez a szerzetes? Először is megfelelő
modellszervezetet keresett, amelynek végül is a borsó, azaz a
Pisum sativum bizonyult. A borsó ugyanis képes önmegtermékenyítésre,
gyorsan nő, kevés helyet igényel, ráadásul, mivel termesztett növény,
rengeteg változata ismert. Mendel harminckét borsótörzsből két
év alatt választotta ki azt a néhány vonalat, amellyel kísérletezni
kezdett. Ezek után hét tulajdonságot választott ki, a további munkája
során ezek öröklődését vizsgálta.
A kiválaszott tulajdonságok:
- borsószem alakja: kerek vagy szögletes;
- sziklevél színe: sárga vagy zöld;
- a virág színe: bíbor vagy fehér;
- hüvely alakja: felfújt vagy szemre simuló;
- hüvely színe: zöld vagy sárga;
- virágok helyzete: axiális vagy terminális;
-
szár hossza: hosszú vagy rövid.
Ezek után az egyes törzseket addig keresztezte, amíg az utódok több nemzedéken át, nagy mintaszám esetén is egyformák voltak, azaz már nem hasadtak (szegregáltak) tovább. Ő ezeket tiszta vonalaknak nevezte, a mai szóhasználattal homozigótáknak hívjuk őket. Egy diploid élőlény ugyanis minden kromoszómájából két példánnyal rendelkezik, az egyiket az egyik, a másikat a másik szülőjétől kapja, így minden egyes génünkből is két példánnyal rendelkezünk. Egy gén különböző változatait alléloknak nevezzük. Mendel a kísérleteiben minden egyes gén két-két allélját vizsgálta. Az általa tiszta vonalaknak nevezett homozigóta növények tehát az adott génnek az egyik allélját örökölték mindkét szülőjüktől, így utódaiknak is csak ezt adhatták át, így a tiszta vonal egyedein belül keresztezve nem hoztak létre más fenotípusú utódokat, azaz az utódnemzedékek nem hasadtak, azaz szegregáltak.
Mendel a kísérleteiben pontosan nyomon követte az egyes egyedek leszármazását, amit úgy végzett, hogy az éretlen borsóvirágokból eltávolította a porzókat, majd egy kis papírtölcsérrel lefedte őket, hogy idegen virágpor ne szennyezhesse a kísérletét. A kísérletében használt tiszta vonalakat, azaz homozigótákat nevezte P, azaz szülői (Parentes) nemzedéknek, míg az ő utódaikat F1, az F1 nemzedék egymás közötti keresztezéséből származó utódait F2 nemzedéknek nevezte el.
Ezek után tiszta vonalú (!)
piros virágú borsót keresztezett tiszta vonalú fehér virágú borsóval
(P – szülői nemzedék). Azt tapasztalta, hogy az utódaik, az F1
nemzedék egyöntetűen piros virágúak lettek. Erre magyarázatot
is adott, a két szülő a virágszínt meghatározó génnek különböző
allélját hordozta, ám mindkettő ugyanabból az allélból kettőt.
Az utódaik mindkét különböző allélből egyet – egyet kaptak,
így egyformák lettek, mivel a piros virágszín domináns a fehér
felett, így a növények fenotípusát ez határozza meg. Gregor Mendel
jelölésével élve: A tiszta vonalú, piros virágú szülő AA, míg
a tiszta vonalú, fehér virágú szülő aa allélokat hordozott. Az
F1 utódnemzedék egyedei az egyik szülőtől egy A allélt kaptak,
mivel nem kaphattak mást, míg a másik szülőtől egy a allélt kaptak,
hiszen tőle sem örökölhettek mást. Így az F1 nemzedék genotípusa
Aa lett. Ez alapján könnyű meghatározni azt is, hogy ugyanannak
a génnek két allélja közül melyik a domináns és melyik a recesszív: domináns az, amely a heterozigóták fenotípusát meghatározza, recesszív
az, amely fenotípusa csak homozigóta formában jelentkezik.
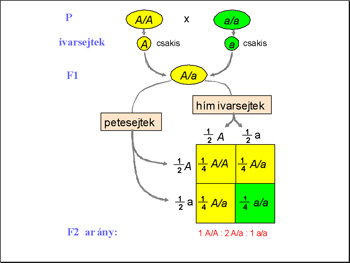 Ezek után az F1 nemzedék
egyedeit egymással keresztezte és meglepődve tapasztalta, hogy a
körükben ismét megjelentek fehér virágú egyedek is. Mendel lángelméje
ott mutatkozott meg, hogy meg is számolta az utódokat, azaz statisztikai
alapon elemezte őket! 705 darab piros virágú és 224 darab fehér
virágú növényt kapott. Ezt az arányt ő 3:1 –nek mérte. Ezek
után kíváncsi volt rá, hogy az utódok vajon tiszta vonalúak –e?
Az F2 nemzedék egyedeit önmegtermékenyítéssel szaporította és
megvizsgálta az utódaikat (F3 nemzedék). Azt tapasztalta, hogy a
domináns fenotípust mutató egyedek harmada tiszta vonalú, azaz ebben
az esetben az utódaik is piros virágúak lesznek, míg a kétharmaduk
nem tiszta vonalú, azaz az utódaik között fehér virágúak is felbukkannak,
míg a fehér virágúak mind tiszta vonalúak. Arra következtetett,
hogy a piros virágúak egyharmada, azaz az összes utód egynegyede
AA genotípusú, azaz tiszta vonalú, kétharmada, azaz az összes utód
fele Aa genotípusú, azaz fenotípusosan piros virágú, ám az utódaik
között fehér virágúak is megjelennek, míg a fehér virágúak
aa genotípusúak, természetesen tiszta vonalúak.
Ezek után az F1 nemzedék
egyedeit egymással keresztezte és meglepődve tapasztalta, hogy a
körükben ismét megjelentek fehér virágú egyedek is. Mendel lángelméje
ott mutatkozott meg, hogy meg is számolta az utódokat, azaz statisztikai
alapon elemezte őket! 705 darab piros virágú és 224 darab fehér
virágú növényt kapott. Ezt az arányt ő 3:1 –nek mérte. Ezek
után kíváncsi volt rá, hogy az utódok vajon tiszta vonalúak –e?
Az F2 nemzedék egyedeit önmegtermékenyítéssel szaporította és
megvizsgálta az utódaikat (F3 nemzedék). Azt tapasztalta, hogy a
domináns fenotípust mutató egyedek harmada tiszta vonalú, azaz ebben
az esetben az utódaik is piros virágúak lesznek, míg a kétharmaduk
nem tiszta vonalú, azaz az utódaik között fehér virágúak is felbukkannak,
míg a fehér virágúak mind tiszta vonalúak. Arra következtetett,
hogy a piros virágúak egyharmada, azaz az összes utód egynegyede
AA genotípusú, azaz tiszta vonalú, kétharmada, azaz az összes utód
fele Aa genotípusú, azaz fenotípusosan piros virágú, ám az utódaik
között fehér virágúak is megjelennek, míg a fehér virágúak
aa genotípusúak, természetesen tiszta vonalúak.
Ezzel a keresztezéssel Mendel "belelátott" az F2 nemzedék piros virágú egyedeinek a genomjába, meg tudta állapítani, mi volt a genotípusuk. Az utódok megoszlása pontosan követte a modell alapján számolt matematikai esélyeket. A piros virágú F1 nemzedék tagjai mind Aa genotípusúak voltak, azaz az utódaik fele az A, másik fele az a allélt örökölte tőlük. Így a modell alapján megjósolható az F2 nemzedék tagjainak a fenotípusa. (Az ábra innen származik.)
Mivel az F2 nemzedék tagjai egyenlő eséllyel öröklik a szülői alléleket, azaz mindkét szülőjüktől 1/2 eséllyel kapják meg az A vagy az a allélt. Tehát ha az első társaságot nézzük, mi az esélye, hogy AA genotípusú egyed jöjjön létre? 1/2 eséllyel örökli az egyik szülőjétől az A allélt, 1/2 eséllyel örökli a másik szülőjétől is az A allélt. Mivel a mindkét esemény véletlennek tekinthető, az együttes bekövetkezésük valószínűségéhez az egyes valószínűségeket össze kell szorozni. 1/2X1/2=1/4, azaz a matematikai modell alapján az F2 nemzedék egynegyede lesz tiszta vonalú piros virágú, azaz AA genotípusú. Ugyanígy kiszámolható, hogy az F2 nemzedék egyedei közül hányan lesznek fehér virágúak, azaz aa genotípusúak: 1/2X1/2=1/4. Ezek után kiszámolható, hogy 1/2 az esélye az Aa genotípus kialakulásának.
Mendel kísérletei pontosan követték a matematikai modellt, azaz tiszta vonalú szülők (P) AA X aa keresztezésekor az F1 utódok egyöntetűen Aa genotípusúak, fenotípusukra nézve a domináns allél által meghatározott fenotípust mutatják, azaz piros virágúak. Az ő beltenyésztésükből származó utódok 1/4 –e AA azaz homozigóta piros virágú, 1/2 –e Aa azaz heterozigóta piros virágú és 1/4 –e aa genotípusú azaz fehér virágú. Azaz az F2 nemzedék fenotípusára nézve 3/4 –ük piros (AA+Aa), 1/4 –ük fehér virágú (aa).
Ezek után megismételte a kísérletet az általa vizsgált tulajdonságokkal és mindegyikkel hasonló eredményt kapott. Ebből arra következtetett, hogy az általa feltárt törvényszerűségek nem egyedi, elszigetelt példák, hanem a növények öröklődésére általánosan jellemzők. Azóta beigazolódott, hogy az általa feltárt folyamatok nem csak a növények öröklődésére igazak, hanem minden diploid szervezetére.
| Szülői fenotípusok | F1 | F2 | F2 arány |
| Piros x fehér virág | mind piros | 705 piros : 224 fehér | 3,15 : 1 |
| Sárga x zöld szem | mind sárga | 6022 sárga : 2001 zöld | 3,01 : 1 |
| Felfújt x befűzött hüvely | mind felfújt | 882 felfújt : 299 befűzött | 2,95 : 1 |
| Zöld x sárga hüvely | mind zöld | 428 zöld : 152 sárga | 2,82 : 1 |
| Axiális x terminális virág | mind axiális | 651 axiális : 207 terminális | 3,14 : 1 |
| Hosszú x rövid szár | mind hosszú | 787 hosszú : 277 rövid | 2,84 : 1 |
Mendel éppen azt igazolta, hogy az egyes allélok öröklődése matematikai esélyeket követ, a recesszív allélok nem lappanganak valamilyen varázslatos módon néhány száz nemzedéken át, hogy aztán látszólag a semmiből újra előtörjenek. Az allélek "újrakeveredése" szigorú statisztikai törvényszerűségeket követ. Két heterozigóta keresztezésekor az utódok negyede a recesszív fenotípust mutatja. Ha egy adott fenotípusért két függetlenül öröklődő gén recesszív allélja a felelős, az utódok 1/16 –oda akkor is a recesszív fenotípust mutatja (1/4X1/4), ha három gén recesszív allélja felelős érte, akkor 1/64 –ede (1/4X1/4X1/4) és így tovább. Az egyes egyedek genotípusa egyszerű keresztezési kísérletekkel vizsgálható, így a "rejtett" recesszív allélek megléte egyértelműen igazolható vagy cáfolható. Mendel éppen azt igazolta, hogy megfelelő kísérleti rendszerben az egyes egyedek genotípusa éppen úgy vizsgálható, mint a fenotípusa.
Vizsgáljuk meg Gregor Mendel
eredményeinek fényében az értelmes tervezés híveinek állításait:
"A legtöbb variáció
a meglévő gének rekombinációjával (újra-elrendeződésével)
keletkezik."
"Az
„új” jelleg, ami látszólag a semmiből bukkan elő, valószínűleg
nem is új, csupán egy recesszív gén kifejeződése, amely mindvégig
jelen volt. Amikor a tenyésztő újfajta kutyát vagy húsosabb marhát
hoz létre, valójában csak a géneket keveri meg egy kicsit, hogy
a recesszív gént kifejezésre juttassa."
Alighanem Mendel eredményeinek
a fényében el kell, hogy vessük azt a feltételezést, hogy évszázadokon
át, több százezer egyed genomjában százszámra lappanghatnának
fel nem ismert recesszív allélok, anélkül, hogy egyetlen homozigóta
recesszív egyed is született volna. Egy allél hiába recesszív,
az utódoknak átadása szigorú matematikai törvényszerűségeket
követ, ezek mérhetőek, százalékos esélyek formájában kifejezhetőek,
ezek alapján az egyes recesszív jellegek fenotípusbeli megjelenése
pontosan megjósolható. Hol vannak a tacskólábú farkasok? Hol vannak
a bulldogpofájú farkasok? Miért nem születnek pincsiméretű farkasok?
Ha ezek az allélek olyan gyakoriak, hogy a tenyésztők "vakon",
azaz a domináns fenotípust mutató egyedek tenyésztésével is mintegy
"véletlenül" begyűjtötték a tenyésztett kutyák közé,
a vadonban miért nem figyelhetőek meg a recesszív allélekre homozigóta
egyedek? Rengeteg farkast tartanak állatkertekben, mégsem születnek
köztük a szerző által feltételezett recesszív jellegeket hordozó
egyedek, pedig, ha ezek a gének ott lappanganának a genomjukban, időnként
mindenképpen kellene születnie egy-egy recesszív fenotípust is mutató
egyednek. Viszont legjobb tudomásom szerint nem írták le a farkasokról,
hogy ilyen elképesztő formagazdagságot mutatnának, márpedig, ha
ezek a recesszív allélek ott lennének a farkas genomban, éppen a
mendeli genetika szabályai szerint kiszámítható eséllyel születniük
kellene a recesszív fenotípusokat mutató egyedeknek. Viszont valamiért
mégis hiányoznak a recesszív allélekre homozigóta egyedek. Hogyan
lehetséges ez? A legvalószínűbb magyarázat annyi, hogy a tenyésztés
során bizony új mutációk is keletkeztek, nem csak a már meglévő
allélek keveredtek újra.
Amúgy érdekes kérdés, hogy
a feltételezett értelmes tervező minek tervezett ennyi fölösleges,
sőt, káros allélt az egyes génekre, ha amúgy sem nyilvánulnak
meg soha a vadállatok fenotípusában? Feltételezem az értelmes tervezés
nem azért történt, hogy az ebtenyésztők egyesületi tagjainak legyen
megfelelő alapanyaga majdan, százezer évekkel később.
"Vegyük szemügyre
a zsiráfot: túlméretezett végtagok, megnyúlt nyak, esetlen testtartás
– látszólag minden ügyetlenül aránytalan. Ennek ellenére a zsiráf
testrészei bámulatos összhangban
állnak egymással: az állat elegáns könnyedséggel mozog,
és olyan erős a rúgása, hogy kevés természetes ellensége van."
A zsiráf nagy méretű, vadon
élő, lassan szaporodó afrikai állat. Szinte mindenki ismeri, viszont
tökéletesen alkalmatlan genetikai modellszervezetnek. Több mint egy
évig vemhes, a nőstény három-négyévesen válik ivaréretté, azaz
egyetlen nemzedékváltást kb. öt év alatt lehetne megfigyelni, ráadásul
rendszerint egyetlen egyet borjadzik, így az utódok statisztikai elemzése
is elég nehézkes. Ennek fényében nem meglepő, hogy tudomásom szerint
még soha senki sem végzett egyetlen genetikai kísérletet sem zsiráfon.
Vitaindítónak viszont tökéletes egy alapvetően genetikai tárgyú
cikkben, hiszen mivel egyetlen sornyi adat sincs az öröklődéséről,
a szabad elméletalkotást nem zavarják holmi tények és kísérleti
eredmények. (Néhány elszórt vizsgálat történt, mikroszatellita
merkereket, mitokondriális DNS-t vizsgáltak, illetve egy interferon
gént, azonban a zsiráf öröklődéséről nincsenek adatok.)
 Nézzünk meg inkább egy példát,
ahol a háziasítás folyamata jól követhető sőt, még az egyes
tulajdonságok öröklődése is ismert. Gondolom nem kell bemutatnom
a szíriai aranyhörcsögöt, amely kedvelt háziállat nálunk is.
Latin neve Mesocricetus auratus. Magyar nyelven is bőséges
forrásanyag áll a rendelkezésünkre róla (pl. itt,
itt, itt, vagy
itt), így nem fáradnék
a bemutatásával.
Nézzünk meg inkább egy példát,
ahol a háziasítás folyamata jól követhető sőt, még az egyes
tulajdonságok öröklődése is ismert. Gondolom nem kell bemutatnom
a szíriai aranyhörcsögöt, amely kedvelt háziállat nálunk is.
Latin neve Mesocricetus auratus. Magyar nyelven is bőséges
forrásanyag áll a rendelkezésünkre róla (pl. itt,
itt, itt, vagy
itt), így nem fáradnék
a bemutatásával.
Miért ilyen különleges ez a rágcsáló? Az első aranyhörcsögöt 1930–ban fogták be, ez egy anyaállat volt az éppen meglévő almával együtt. A fiatalok közül mindössze három egyedet, egy nőstényt és két hímet tenyésztettek tovább, a ma fogságban élő aranyhörcsögök ettől a három egyedtől származnak (Később alapítottak néhány más tenyészállományt is, újabb befogott egyedekből, de ezekkel a populációkkal nem foglalkoznék.). A vad típusú hörcsög – meglepő módon - aranyszínű bundájú, ám az idők során a tenyésztők több különböző színváltozatot alakítottak ki. Ezek közül csak néhányat tárgyalnék, természetesen a teljesség igénye nélkül.
Létezik egy halványszürke színváltozat, amelyet az lg gén domináns allélja (Lg) okoz. Ismert egy ezüstszürke változat, amelyet az sg gén domináns allélja (Sg) okoz. A sárga színt az X kromoszómához kötött to gén domináns alléja (To) alakítja ki. Ezeken kívül léteznek a bunda mintázatát módosító gének is. A ragyogó szatén színt az sa gén domináns allélja (Sa) okozza. Ezeken kívül domináns jelleg a háton végigfutó fehér csík (ba gén Ba allélja), illetve a bunda fehér foltossága (ds gén Ds allélja), valamint a fehér has is (wh gén Wh allélja). Mivel az összes szelídített aranyhörcsög attól a nyolcvan éve élt három jószágtól származik, kijelenthetjük, hogy ezek az allélok bizony mind a szelídítés során jelentek meg, új mutációs események nyomán, egyszerűen azért, mert dominánsan öröklődnek, azaz az alapító egyedek nem hordozhatták őket a genomjukban észrevétlenül, mindenképpen megjelent volna a fenotípusukban is, azaz látni kellett volna, ha ezeket az allélokat hordozzák. Az aranyhörcsög esetében kizárhatjuk, hogy vad populációkkal történt volna valamilyen keveredés, hiszen fogságban tenyésztik, ráadásul vadon csak a világ egy szűk területén él. Ezzel a rövidke példával azt hiszem a boncolgatott cikk minden kiindulási állítását sikerült cáfolnom, egy megfelelő modellállat segítségével:
- Ezek az allélek dominánsak, azaz nem minden, a tenyésztés során megjelenő tulajdonság öröklődik recesszíven.
- Ezek az allélek a populáció alapítóegyedeiből biztosan hiányoztak, ha meglettek volna, a fenotípusban is jelentkezniük kellett volna, tehát szükségképpen új mutációs események során alakultak ki.
- A tenyésztés során láthatólag nem csak a recesszív allélok újrakeverése történt, hanem bizony új domináns (!) allélok is megjelentek.
"Ezzel szemben a mendeli
modellben a gének sokkal inkább különálló
részecskékhez hasonlítanak, amelyek az
öröklődés folyamán nem változnak."
" Elfogadható becslés, ha azt mondjuk, hogy egy 1000 fős populációban
egyetlen egyedben egyetlen
mutáció fordul elő."
"Hogyan megy végbe
a változás a mendeli genetika keretein belül?"
Drake J. W. és munkatársai
1998–as cikkében találunk néhány érdekes adatot az egyes szervezetekben
megfigyelt spontán, azaz minden külső beavatkozás nélkül bekövetkező
mutációk gyakoriságáról. A spontán mutációk bekövetkezésének
esélye egy szervezeten belül az egyes gének esetében sem egyforma,
például kukoricában a spontán mutációk bekövetkezésének esélye
a több gén átlagában: 7,7X10-5 . Ennek fényében az
1X10-3 becslés, amit a szerző hoz éppen csak százszor-ezerszer
több, mint a valóságban bekövetkező mutációk száma. Ezzel meg
is fejtettük az okát, miért bizonyultak olyan állandónak a gének
Mendel kezeiben: Ő a kísérleteiben néhány száz növényt vizsgált
végig, a legnagyobb egyedszámot is a borsószemek vizsgálatakor elemezte,
körülbelül nyolcezer egyedet. Kukoricában átlagosan körülbelül
minden hétszázhetvenezredik egyedben történik mutáció egy adott
génben, tehát matematikai esélye akkor lett volna, hogy Mendel egyetlen
egy mutációs eseményt megfigyeljen a borsószem színében, ha körülbelül
százszor ennyi egyedet vizsgált volna a kísérletben, a többi tulajdonság
vizsgálatakor pedig még ennél is kisebb mintaszámmal dolgozott.
Ezek alapján elég könnyű megfejteni ezt a rejtélyt: Spontán mutációk
lényegesen ritkábban történnek, mintsem hogy Mendelnek esélye lett
volna meglátni őket a néhány száz egyeddel végzett kísérleteiben,
így lehet, hogy ő a géneket megváltoztathatatlan egységnek látta.
Egyszerűen túl kevés egyedet vizsgált ahhoz, hogy mutációs eseményeket
figyelhessen meg. Ugyanezen cikk szerint egy egérben 6,6X10-6
ecetmuslicában 8X10-6 eséllyel történnek mutációk.
Ez alapján akárhogyan is nézzük, a szerző becslése lényegesen
több mutációval számol, mint amennyi a valóságban megtörténik.
" Továbbá
a beltenyésztés a hibás jellegek koncentrációját eredményezheti,
és ahogy a faj átlagos morfológiájától egyre
távolabb kerülünk, egyre több disszonanciát, stresszt
és csökkenő termékenységet tapasztalunk.
Az ilyen túltenyésztett populáció
gyakran hajlamos visszafejlődni a faj
átlagos morfológiai irányába."
Ha a szerző maga szögezi
le, hogy új allélok nem keletkeznek, csak a meglévőek eloszlása
változik, hogyan fejlődhetnének vissza a beltenyésztett populációk?
A szerző – nem mellesleg teljesen alaptalan - állítása szerint
minden egyes tenyésztett állatban meglévő minden egyes jelleg recesszíven
öröklődik, a tenyésztők mindössze ezeket a recesszív alléleket
hordozó egyedekből hoznak létre tiszta vonalakat, azaz állítják
homozigóta állapotba. Amennyiben az adott, túltenyésztett jelleget
okozó tulajdonság recesszíven öröklődik, akkor nem változhat
meg mutációs esemény nélkül, hiszen az egyed homozigóta, azaz
"aa" genotípusú, onnan tudjuk biztosan, mert különben
a domináns fenotípust mutatná. Ebben az esetben teljesen érthetetlen,
hogyan "hajlamos az ilyen túltenyésztett populáció
visszafejlődni a faj átlagos morfológiai irányába", hiszen
ehhez az egyedeknek valahonnan varázslatos módon génenként egy domináns
allélt kéne szerezniük a semmiből, hiszen az ő genomjukban ez nem
lehet meg. Hogyan lehetséges ez? Mert ugye ha ez bekövetkezne, akkor
éppen az a hasznos mutáció lenne, amely az értelmes tervezés hívei
szerint nem is létezik.
" A mutációk létrehozhatnak
például összegyűrődött, túl- vagy
alulméretezett szárnyakat. Eredményezhetnek megduplázódott szárnyakat
is, de nem eredményeznek újfajta szárnyakat,
és főleg nem alakítják át a gyümölcslegyet egy
újfajta rovarrá."
" Nincs azonban az
a mennyiségű finombeállítás, ami a jelenlegi testfelépítéséből
egy teljesen új testfelépítést tudna létrehozni."
Az ilyen kijelentések előtt
érdemes lenne tisztázni, mi számít "újfajta" szárnynak?
Ha az nem, ami nagyobb, az sem ami kisebb, az sem amiből kettő helyett
négy van? Nem tudom, nekem az, hogyha a légynek két szárnya helyett
négy nő, eléggé új testfelépítés. Például ha egy ember négy
karral születne, vagy a kézfejei közvetlenül a vállából állnának
ki, esetleg a karjai leérnének a földig, vagy a karjai helyett is
lábai nőnének, nem mondanám rá, hogy ez teljesen olyan, mint a
többi, semmi új nincs benne. Sokkal de sokkal könnyebb lenne érdemi
vitát folytatni, ha az értelmes tervezés hívei meghatároznák az
elvárásaikat. Ha eldöntenék, hogy mi számít "újfajtának",
akkor lehetne rá példát hozni. A jelenlegi helyzet az, hogy az "újfajta"
meghatározása annyi: Amilyen mutációt valaha is leírtak, az éppen
nem "újfajta". Mikor lenne elfogadható az eredmény, azaz
mik az "újfajta" követelményei? Sajnos erre semmilyen elfogadható
meghatározást sem kapunk, ami, ha meggondoljuk valóban kényelmes,
hiszen így akármilyen példát hozna is egy biológus, arra rá lehet
mondani, hogy pont nem az.
"Másképpen fogalmazva,
a tenyésztők édesebb kukoricát, húsosabb tehenet tudnak produkálni,
de nem tudják a kukoricát más növénnyé
vagy a marhát más állattá alakítani."
Érdemes megvizsgálni a Raphanobrassica
nevű növényt. Ezt
Gregorij Karpecsenkó szovjet tudós hozta létre 1928–ban, tehát
nem igazán új eredmény. Retek és káposzta növényeket keresztezett,
ám úgy, hogy mindkettőt kolhicinnel kezelte, így a kromoszómák
nem váltak szét az ivarsejtképzéskor és az utódba mindkét növény
teljes genomja bekerült. A retek és a káposzta diploid kromoszómaszáma
egyaránt 18, az új növény diploid sejtjeiben 36 kromoszóma található.
Amiért érdekes: Ez a növény egyáltalán nem terméketlen, ivarsejteket
képez, magot hoz, így minden további nélkül termeszthető, azonban
egyik szülői fajjal sem képes termékeny utódot létrehozni. Sajnos
a növény gyökere a káposztáéra, levele pedig a retekére hasonlít,
így nem igazán ehető, de kétségkívül ez egy olyan eset, amikor
a tenyésztők két fajt egy teljesen más fajjá alakítottak, amelyik
külső-belső tulajdonságaiban különbözik mindkét szülői fajtól.
Másik, később végzett növénynemesítési munka a Triticale, azaz
a búza és a rozs teljes genomját hordozó gabona megalkotása, az
ötvenes években. A termesztett búza diploid kromoszómaszáma 42,
a rozsé 14, nem csoda, hogy a Triticale diploid kromoszómaszáma 56.
A Triticale mindkét szülői faj jellegzetességeit magán viseli.
A tenyésztők igenis képesek egyes növényeket "más növénnyé"
alakítani.
"Ezzel szemben a mendeli
modellben a gének sokkal inkább különálló
részecskékhez hasonlítanak, amelyek az
öröklődés folyamán nem változnak."
"A mutációk szerencsére eléggé ritkák, mert túlnyomó többségük ártalmas, és csak néhány
semleges."
"Ugyanígy a
mutációk is csak elvétve javítják a DNS
üzenet minőségét, és túl sűrű
előfordulásuk akár halálos is lehet a szervezetre nézve."
"A gének rekombinációjával
ill. mutációkkal generált kicsiny változások elősegíthetik ugyan
a természetes kiválasztódás által a szervezet
„finombeállítását”, lehetővé
téve, hogy ökológiai környezetéhez
jobban alkalmazkodjék."
"Ezek a behatárolt
változások nem akkumulálódnak, ahogy a darwini evolúciós teória
megkívánná az új makrováltozások létrehozásához."
Ha a cikk elején azt állítja,
hogy Mendel igazolta a gének állandóságát, miért ír folyton mutációkról?
Most akkor mutációk egyáltalán léteznek, vagy nem? Már csak azért,
mert a mutációk létét elég nehéz lenne tagadni. Majd a szerző
maga is elismeri, hogy léteznek előnyös mutációk, amelyek "javítják
a DNS üzenet minőségét", ám néhány sorral feljebb határozottan
állítja, hogy mindegyik mutáció káros, vagy semleges. Akkor most
léteznek előnyös mutációk, vagy nem? Ha a mutációk elősegíthetik
a szervezet "finombeállítását", akkor ezek szerint mégis
vannak előnyös mutációk. Ha állítja, hogy az egyes előnyös mutációk
nem halmozódnak fel, miért beszél éppen a "gének" "finombeállításáról",
így többes számban? Ezek szerint akkor a hasznos mutációk mégis
csak felhalmozódhatnak? Az előnyös mutációk felhalmozódása nem
éppen az az evolúció, ami a szerző szerint nem is létezik? Miért
ennyi az önellentmondás ebben az alig néhány oldalas rövid szövegben?
Mi a legnagyobb bajom ezzel
a cikkel? Az, hogy egyetlen egy adat sincs benne. Ír a zsiráfról,
amely állat genetikáját soha senki sem vizsgálta, valamint a kutyákról,
amelynek öröklődése elég kevéssé tanulmányozott, de egyetlen
adatot sem hoz a kutya génjeiről sem. Egyetlen tulajdonság öröklődéséről
sem ír, csak homályos általánosságokat említ. Ráadásul a cikk
állításai minden alapot nélkülöznek, egyszerűen vonzó közhelyek.
Miért van az, hogy az értelmes tervezés hívei, akik az evolúciós
elméletek elemzésekor a legapróbb részletekre is kísérletes bizonyítékot
követelnek, a saját elméleteik esetében ettől nagyvonalúan eltekintenek?
A szerző teljesen alaptalan tényeket szögez le, majd ezekből von
le messzemenő következtetéseket. Mint bölcseleti munka vagy szépirodalmi
alkotás talán megállja a helyét, de semmilyen kapcsolatban sincs
a valósággal. Olvassuk el, mit is jelent pontosan a tudományos módszertan!
- lépés: Megfigyelés és
kísérletezés - Adatgyűjtés
-
lépés: Feltételezés
– Az adatok magyarázata
-
lépés: Előrejelzés –
A feltételezés alapján a rendszer működésére és jövőbeli változásaira
vonatkozó "jóslatok" megtétele.
-
lépés: Ellenőrzés –
A feltételezés talaján álló előrejelzések összevetése a valósággal.
Az éppen vizsgált cikk a
tudományos módszer melyik lépését tartja be? Teljesen alaptalan,
egymásnak részben ellentmondó feltételezésekből és minden kísérleti
bizonyítékot nélkülöző kijelentésekből indul ki. Adatgyűjtést
nem végez. Feltételezéseit valóban kibontja, ám ezek adatok híján
teljesen értékelhetetlenek, csak lógnak a levegőben. Előrejelzéseket
nem tesz, és nem ellenőrzi, hogy az általa felépített feltételezések
milyen viszonyban állnak a valósággal. Ahol az állításai ellenőrizhetőek,
azaz egy megfelelő modellrendszerben, például a szíriai aranyhörcsög
esetében, ellent is mondanak a természetben megfigyelt tényeknek,
azaz az ellenőrzés során elbuknak. Ki kell hogy jelentsük, hogy
tetszetős elmélet, ám a valósághoz semmi köze sincs.
Drake J. W., Charlesworth B., Charlesworth D., Crow J. F. (1998) Rates of spontaneous mutation. Genetics 148(4): 1667-1686.