


 Ha egyszer majd valaki veszi a fáradtságot, hogy összeállítsa, a legfélrevezetőbb magyar állatnevek listáját, a lándzsahal mindenképpen előkellő helyre fog befutni. Ugyanis ez a kis állat nem hal, de mégcsak nem is gerinces. Ettől persze a hasonlóság még valós, de közel sem annyira szoros rokonságot takar (mint látni fogjuk, sőt), mint azt a nevezéktan sugallja.
Ha egyszer majd valaki veszi a fáradtságot, hogy összeállítsa, a legfélrevezetőbb magyar állatnevek listáját, a lándzsahal mindenképpen előkellő helyre fog befutni. Ugyanis ez a kis állat nem hal, de mégcsak nem is gerinces. Ettől persze a hasonlóság még valós, de közel sem annyira szoros rokonságot takar (mint látni fogjuk, sőt), mint azt a nevezéktan sugallja.
A lándzsahal, vagy régiesebb nevén amphioxus, ugyanis a fejgerinchúrosok (Cephalochordata) közé tartozik. Ezek az állatok néhány nagyon fontos közös jellegben osztoznak, a saját magunk által is tarkított gerincesekkel, különösen az embrionális jellegeket tekintve. Ilyenek a kopoltyúívek, a test felső/hátsó oldalán végigfutó központi idegrendszer, és az alatta megtalálható pálcika-szerű gerinchúr. Persze legalább annyira fontosak a különbségek is, pl. a jól fejlett fej, porcok és csontok ill. végtagok teljes hiánya (hogy csak a legszembetűnőbbeket említsem), de igazából a poszt szempontjából ez egy kicsit mellékvágány, így elegánsan eltekintünk további boncolgatásuktól.
Az említett hasonlóságok miatt a lándzsahalak mindenesetre már bő száz éve erőteljes érdeklődésre tartanak számot biológusi körökben, gyakran számos feltüzelt vita kiindulópontját képezve. Például 1910 környékén a kor két jeles brit zoológusa, Walter Gaskell és Edwin Goodrich feszült egymásnak, azt eldöntendő, hogy az amphioxus egy degenerált gerinces, vagy épp ellenkezőleg, a gerincesek ősi vonásait megjelenítő "primitív" faj. Goodrich-nek lett igaza, de talán még ő maga sem sejtette, hogy a lándzsahalk ősisége mennyire régre vezethető vissza: az elmúlt években előkerült fosszilis anyag tanúsága szerint már több mint bő félmilliárd éve nagyon hasonló testfelépítésű állatok szelték a kambriumi óceánok vizeit.
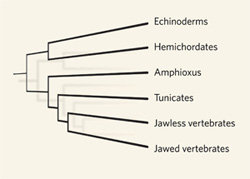 A gerincesek és fejgerinchúrosok útjai durván 550 millió éve váltak el és a közkeletű vélekedéssel ellentétben (ami ma még gyakran tankönyvi anyag), az utóbbi évek molekuláris rendszertana kiderítette, hogy a két csoport ettől még nem a legközvetlenebb rokon az Újszájúak szupertörzsében. Ugyanis a jelek szerint a gerincesek felé vezető útból még később leválik a mára mind alakjukban, mind genom szerkezetükben rendkívül módosult zsákállatokat (Tunicates) magába foglaló előgerinchúrosok vonala. Őket azonban épp ez az extrém módosulás teszi alkalmatlanná arra, hogy a gerincesek ősienek titkait kutassuk bennük, erre viszont ideális a rokonsági sorban paraszthajszálnyit hátrébb csúszó kis lándzsahal.
A gerincesek és fejgerinchúrosok útjai durván 550 millió éve váltak el és a közkeletű vélekedéssel ellentétben (ami ma még gyakran tankönyvi anyag), az utóbbi évek molekuláris rendszertana kiderítette, hogy a két csoport ettől még nem a legközvetlenebb rokon az Újszájúak szupertörzsében. Ugyanis a jelek szerint a gerincesek felé vezető útból még később leválik a mára mind alakjukban, mind genom szerkezetükben rendkívül módosult zsákállatokat (Tunicates) magába foglaló előgerinchúrosok vonala. Őket azonban épp ez az extrém módosulás teszi alkalmatlanná arra, hogy a gerincesek ősienek titkait kutassuk bennük, erre viszont ideális a rokonsági sorban paraszthajszálnyit hátrébb csúszó kis lándzsahal.
A lándzsahalak 19 kormoszóma párt elfoglaló ~520 megabázisnyi genomja lényegesen kisebb, mint az emberi faj maga hárommilliárd bázispárja, bár természetesen nem a méreten van a hangsúly (mert akkor aligha lehetne egy teljes fugu halat "mindössze" 365 millió bázispárból "összehozni"). Viszont a méretbeli eltérés egyik lehetséges oka ez esetben, hogy jelen tudásunk szerint (amit a lándzsahal genom csak megerősített), a fejgerinchúrosoktól való elválás óta minimum két teljes genomduplikáció következett be a gerincesek elődeiben. Azaz minden génből elméletileg kb. négy kellene legyen, s hogy ez mégsincs (egyébként megközelítőleg sem) így, az elsősorban annak köszönhető, hogy a redundáns ill. funkció nélküli gének gyakorta kiszóródnak az idő során, fosszilizálódnak, majd teljesen degenerálódnak.
A genomduplikációk eredménye például az emlősökben fellelhető négy db. Hox-kluszter is, amelyek a testfejlődés számára kulcsfontosságú géneket hordoznak. Ezek közül egyik sem teljes, mindegyikből más gének hiányoznak. Viszont összevetve őket, látható, hogy a gerincesek valamelyik ősének, minimum 13 hox gént kellett felsorakoztatnia egy kromoszómáján. És a lándzsahalban pont egy ilyen szituációval találkozunk (sőt, itt egyenesen tizenöt hox gén lelhető fel; nem teljesen tisztázott még, hogy a közös ősben is ennyi volt, csak a gerinces vonal még a genomduplikációk előtt elvesztett kettőt, vagy az amphioxus "ajándékozta meg" még kettővel magát).
Egyébként, ha pusztán azt nézzük, a fent említett Hox-kluszter mintájára, hogy milyen gyakran találjuk kb. ugyanazokat a géneket egymáshoz közel a lándzsahal és az ember kromoszómáján (ez szaknyelven a szinténia jelensége, aminek az oka, hogy - értelemszerűen - a két faj közös ősében ezek a gének szintén szomszédosan helyezkedtek el), akkor azzal szembesülhetünk, hogy az amphioxus genomjának közel 88%-a esik ebbe a kategóriába (ráadásul, a már emlegetett genomduplikációk miatt, négyszeres szinténia sejlik fel). Na persze a hidrák genomja után, ez nem túl meglepő, mint ahogy az sem, hogy a gének fehérjekódoló szakaszai (szakzsargonban exonok) közé beékelődő DNS darabok, vagyis intronok, a hasonló génekben többnyire pont ugyanott helyezkednek el (másképp fogalmazva, mind a gerincesek, mind a fejgerinchúrosok exon-intron szerkezete meglehetősen konzervatív).
Természetesen ma már nincs genomvizsgálat az ún. konzervált nem-kódoló szekvenciadarabok, CNE-k (Conserved Non-coding Elements) feltérképezése nélkül. Ezek olyan rövid DNS szakaszok, amelyek nem esnek ugyan a genom fehérjekódoló részeibe, mégis nagyon hasonlítanak sok, gyakran egész távoli rokonságban levő faj között. Funkciójuk leginkább a génszabályozással hozható összefüggésbe, pontosabban azzal, hogy egy-egy gén hol kerül kifejeződésre. Minnél hasonlóbb CNE kombináció lelhető fel két faj egy bizonyos génje előtt, annál valószínűbb, hogy az pont ugyanazokban a szervekben, szövetekben kerül kifejeződésre mindkét szervezetben. Míg halak és emberek között ezrével fedezhetők fel a CNE-k, addig a lándzsahal és az emlősök között alig néhány tucatra lelhetünk.
A különbség azoknak az evolúciós "újjításoknak" a számlájára írható, amelyek a gerincesek kialakulásának hajnalára tehetők. Ezek közül is talán a legjelentősebb egy igazi sejtszintű "svájcibicska", az idegrendszer és a kültakaró embrionális előfutárjainak határán kialakuló ún. velőléc (neural crest) sejtpopuláció. Ezeknek a sejteknek a fejlődési potenciálja már-már megdöbbentő: belőlük jön létre koponyánk és nyakunk porcos és csontos elemeinek többsége, a szimpatikus és paraszimpatikus idegrendszer, a mellékvesénk jelentős része, na és a pigment sejtek. Szóval nem nagyon tudnánk őket nélkülözni, s ha valami fejlődési rendellenesség miatt sérül a kialakulásuk, akkor gebasz van. Fontosságuk miatt rengetegen kutatják őket, így egyre többet tudunk arról, hogy miként és hogyan alakulnak ki.
Érdekesség, hogy a jelek szerint a lándzsahalakban az összes gén megtalálható, ami a gerincesekben a velőléc sejtek kialakulásához szükséges genetikai hálózatban szerepel, sőt, a hálózat kezdeti elemei gyakorlatilag épp ott és épp úgy fejeződnek ki (és működnek együtt), mint pl. egy átlagos hal embrióban. A különbség az utolsó lépésekben van: az amphioxus nem "drótozta" be ebbe a hálóba azokat a géneket, amelyek a velőléc sejtek specifikációjának utolsó lépésit irányítanák. Például ilyen a mitf gén, ami bennünk a pigmentsejtek fejlődéséért felel. A lándzsahalban ez a gén mindössze egy apró sejtcsoportban fejeződik ki, ami nem is a leendő idegrendszer és köztakaró sejtjeinek határán található. Viszont ez a pár sejt, az ún. Hesse sejtek, a felnőtt állatban pigmentet fog létrehozni, vagyis a mitf gén aktiválása és a pigment szintézis közti összefüggés létezik. A mitf szerepe valami hasonló lehetett a gerincesek ősében is, még végül az evolúció szeszélyének köszönhetően, a megfelelő szabályozó mutációk megjelenése után bekapcsolódott a (leendő) velőléc-szabályozó hálózatba. S mivel szerepe hasznosnak bizonyult, azóta ott is maradt.
Putnam NH, Butts T, Ferrier DE, Furlong RF, Hellsten U, et al. (2008) The amphioxus genome and the evolution of the chordate karyotype. Nature 453: 1064-1071.
Holland LZ, Albalat R, Azumi K, Benito-Gutiérrez E, Blow MJ, et al. (2008) The amphioxus genome illuminates vertebrate origins and cephalochordate biology. Genome Res doi: 10.1101/gr.073676.107
Yu JK, Meulemans D, McKeown SJ, Bronner-Fraser M (2008) Insights from the amphioxus genome on the origin of vertebrate neural crest. Genome Res doi: 10.1101/gr.076208.108.


{kind=link}