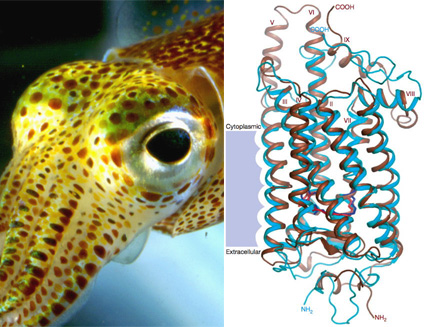
 A felépítés logikája gyakorlatilag ugyanaz és az érzékelés alapegysége sem különbözik nagyon. Az emlősök szeme és egy tintahalszem közt felfedezhető hasonlóság első pillantásra egészen lenyűgözőnek tűnhet, hiszen nemcsak mindkét esetben ún. "kamera" szemről van szó (ahol a fény egy lencsén keresztül jutva vetül a fényérzékeny sejtekre), de mint azt a jobboldali ábra mutatja, a fénydetektálást végző molekula alakja is kísértetiesen hasonló (a kék szín egy szarvasmarha opszin molekulájának tér szerkezetét jelöli, a barna pedig a nemrég publikált tintahal opszin fehérjeszerkezetét). Sőt, lenyűgöző módon, a szemfejlődést elindító gének is gyakorlatilag ugyanazok.
A felépítés logikája gyakorlatilag ugyanaz és az érzékelés alapegysége sem különbözik nagyon. Az emlősök szeme és egy tintahalszem közt felfedezhető hasonlóság első pillantásra egészen lenyűgözőnek tűnhet, hiszen nemcsak mindkét esetben ún. "kamera" szemről van szó (ahol a fény egy lencsén keresztül jutva vetül a fényérzékeny sejtekre), de mint azt a jobboldali ábra mutatja, a fénydetektálást végző molekula alakja is kísértetiesen hasonló (a kék szín egy szarvasmarha opszin molekulájának tér szerkezetét jelöli, a barna pedig a nemrég publikált tintahal opszin fehérjeszerkezetét). Sőt, lenyűgöző módon, a szemfejlődést elindító gének is gyakorlatilag ugyanazok.
Mielőtt azonban egyáltalán komolyabban megfogalmazódna bennünk, hogy netán mindez egy égi fiókban porosodó tervrajz létét igazolná, nem árt egy második pillantást is vetnünk vizsgálataink tárgyára, hogy feltűnjön a köztük levő (számos) eltérés is.
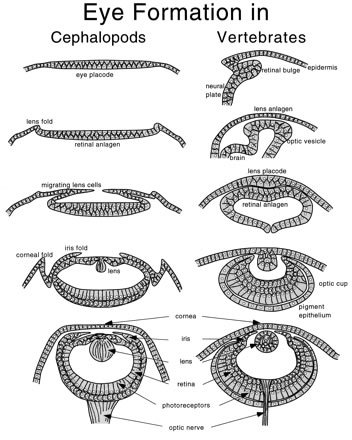 Mindenekelőtt, az emlős retinával ellentétben, a tintahalak esetében a fénynek nem kell átverekednie magát ereken és idegsejtek sűrű sorain, hogy a fényérzékeny fotoreceptorokhoz jusson, hanem közvetlenül azokra vetül, mert a puhatestűek retinájában ezek helyezkednek el "legfelül" (már ha a "fent" ez esetben a lencséhez közelt jelent). Persze, nemcsak a receptorok helyzete, de már alakjuk is jelentősen eltérő, hiába használja minkét csoport az opszin molekulákat a fény érzékeléséhez. Míg az egerek, emberek és tsaik szemében ún. "ciliáris" fotoreceptorok vannak (a pálcika- és csapsejtek), ahol az opszinmolekulák a sejtek ostorának egy különleges módosulatán találhatók, addig a tintahalakban "rhabdomerikus" sejtekkel találkozunk. Itt az ostor nem módosult, az opszinok pedig a sejtek felszínén levő mikrobolyhokban sorakoznak (itt egy részletesebb összehasonlítás).
Mindenekelőtt, az emlős retinával ellentétben, a tintahalak esetében a fénynek nem kell átverekednie magát ereken és idegsejtek sűrű sorain, hogy a fényérzékeny fotoreceptorokhoz jusson, hanem közvetlenül azokra vetül, mert a puhatestűek retinájában ezek helyezkednek el "legfelül" (már ha a "fent" ez esetben a lencséhez közelt jelent). Persze, nemcsak a receptorok helyzete, de már alakjuk is jelentősen eltérő, hiába használja minkét csoport az opszin molekulákat a fény érzékeléséhez. Míg az egerek, emberek és tsaik szemében ún. "ciliáris" fotoreceptorok vannak (a pálcika- és csapsejtek), ahol az opszinmolekulák a sejtek ostorának egy különleges módosulatán találhatók, addig a tintahalakban "rhabdomerikus" sejtekkel találkozunk. Itt az ostor nem módosult, az opszinok pedig a sejtek felszínén levő mikrobolyhokban sorakoznak (itt egy részletesebb összehasonlítás).
Ezek után talán már nem meglepő, ha azt is elárulom, hogy a két szemtípus is nagyon különbőző módon jön létre: míg a gerincesek szeme az idegrendszer, pontosabban a velőcső egy kitüremkedéséből alakul ki (kivéve a lencsét és a szaruhártyát), addig a fejlábúak esetében a teljes struktúra egy felszíni szövetvastagodásból fog kialakulni.
És bizony a működésben is alapvető különbségek vannak: a gerinces szem fotoreceptorai ugyanis sötétben aktívak és fényben "hallgatnak", a tintahal receptorok pedig vice versa. A különbség oka, azokban a molekulákban keresendő, amelyeknek az opszin-molekulák átadják az információt. Ezek ugyan mindkét esetben az ún. "G-fehérjék" osztályába kapcsolódnak, de a hasonlóság itt meg is áll: míg a gerinceseknél a folyamat végeredménye a sejt falában levő (előzőleg aktív) ioncsatornák bezáródása lesz (ezért "hallgat" el, az addig aktív sejt), a tintahalaknál épp ellenkezőlég, megnyílnak az (előzőleg inaktív) ioncsatornák, a sejt pedig tüzelni kezd.
Magyarán, minden hasonlóságuk ellenére, a két szemtípus közt nincs mélyebb közeli rokonság; létük a konvergens evolúció klasszikus példája. S hogy akkor mire véljük az elején megemlített fejlődésgenetikai hasonlóságokat? Nos, a homológ gének jelenléte mindössze arra bizonyíték, hogy a réges-régen élt közös ős feltehetően már rendelkezett egy fényérzékeny szervvel*, amelyből aztán év százmilliók során, nagyon különböző átmeneteken keresztül végül valami hasonló alakult ki. Hasonló, de nagyon nem ugyanaz.
* A szakmabeli kánon szerint ebben az ősi szervezetben valamilyen formában minkét fotoreceptortípus - mind a ciliáris, mind a rhabdomerikus - jelen volt, de az evolúciós fa különböző ágain a szelekció és végül különböző receptorokat juttatott domináns helyzetbe a fényérzékeny szervekben. Ezt látszik igazolni, hogy egyes gerinctelen fajokban még mindkét sejttípus fellelhető, ill., hogy bennünk emlősökben is megtalálható egy, a rhabdomerikus fotoreceptorokra jellemző opszin molekula(-szerűség). Ez a melanopszin és a bioritmus kialakításáért felelős.
Murakami M, Kouyama T (2008) Crystal structure of squid rhodopsin. Nature 453: 363-369.