A második világháború után, a japán frontról visszatérő Ed Lewis újult erővel vetette bele magát a Drosophila genetikában, a Caltechen levő laborjában. Miközben a homeotikus mutációkat vizsgáló, később Nobel díjjal honorált projektjén dolgozott egy érdekes problémával találta magát szemközt: ha két, látszólag egyforma (vagy nagyon-nagyon hasonló) mutánsom van, akkor miképp állapíthatom meg, hogy a mutációk azonos génben vannak, vagy csak ugyanazon folyamat két, különböző génjében. A probléma megoldása végett fejlesztette ki Lewis az ún. genetikai komplementációt, amelyet logikai egyszerűsége és eleganciája azóta is a genetikusok kedvelt kísérletévé tesz.
A második világháború után, a japán frontról visszatérő Ed Lewis újult erővel vetette bele magát a Drosophila genetikában, a Caltechen levő laborjában. Miközben a homeotikus mutációkat vizsgáló, később Nobel díjjal honorált projektjén dolgozott egy érdekes problémával találta magát szemközt: ha két, látszólag egyforma (vagy nagyon-nagyon hasonló) mutánsom van, akkor miképp állapíthatom meg, hogy a mutációk azonos génben vannak, vagy csak ugyanazon folyamat két, különböző génjében. A probléma megoldása végett fejlesztette ki Lewis az ún. genetikai komplementációt, amelyet logikai egyszerűsége és eleganciája azóta is a genetikusok kedvelt kísérletévé tesz.
Az elv tényleg triviális: ha adott A és B mutáns, amelyeket egy-egy recesszív mutáció okoz (vagyis a mutánsok csak akkor szúrhatók ki, ha az adott génből a homológ kromoszómapárjaikon levő mindkét másolat hordozza a mutációt) egyszerűen keresztezem őket és azt figyelem, hogy utódjaik között látható-e a szóbanforgó elváltozás. Ha nem, akkor a két mutáció különböző génekben volt jelen, s mivel az utódoknak csak az egyik szülőtől származó kromoszómáján volt jelen a mutáns gén (mindkét esetben), azaz heterozigóták, ők maguk kinézetre nem lesznek mutánsok. Ellenben ha a mutáns kinézet jelen lesz az utódok közt, akkor A és B mutáció ugyanabban a génben van jelen.
Lewis esetében a mutációk olyan génekben voltak jelen, amelyek a muslicák szárny mögött található egyensúlyozó szervét, billérét "alakították át" valódi szárnnyá, de értelemszerű, hogy a komplementációs analízis működik szinte bármilyen állatban, szinte bármely mutáció esetében. Például egy mexikói lazacfaj, az Astyanax szemnélküliséget okozó mutációi esetében.
Az Astyanax-nak számos populációja él a Yucatán félszigetet ementálihoz hasonlóan felszabdaló mészkő barlangokban és ezeknek a halaknak a többsége vak. Hogy pontosan miért is (mármint elvi szinten miért), arról nincs teljes egyetértés, három nézet is népszerű, és ma még nem tudjuk eldönteni, hogy melyik is áll közelebb a valósághoz. Az első iskola úgy véli, egyszerűen arról van szó, hogy a sötétben a halak nem használják a szemüket, így igazából csak idő kérdése volt, hogy egy szemsorvasztó mutáció megjelenjen és elterjedjen (hiszen nincs szelekció ellene). Egy másik társaság szerint a szem elvesztése kifejezetten előnyös volt, ui. a retinában levő fotoreceptor sejteknek nagy az energiaigénye, márpedig a barland az egy táplálékban szegényes környezet, vagyis minden kis energia megtakarítás előnyt jelent a túlélésben. A harmadik elmélet hívei úgy látják, hogy a hal barlangi élethez való adaptációja során következtek be olyan mutációk, amelyek a fittség növelése mellett, mintegy mellékesen okozták a szemek elvesztését - őket látszik igazolni az a megfigyelés, hogy a barlangi halakra jellemző nagyobb állkapocs ugyanazokkal a génexpressziós változásokkal hozható kapcsolatba, mint a vakság.
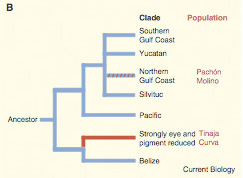 Az elvi részen túl persze még azt sem tudjuk pontosan, hogy az egyes halakban milyen mutáció okozza a szem elsorvadását, mindenesetre mivel számos vak Astyanax populáció nagyon távol él egymsától, feltételezhető, hogy a párhuzamos konvergens evolúció klasszikus példájával állunk szemben, vagyis a vakság egymástól függetlenül alakult ki. Ennek tesztelésére tökéletesen alkalmas a komplementáció, épp ezért végezte ezt el az NYU egyik kutatója, Richard Borowsky.
Az elvi részen túl persze még azt sem tudjuk pontosan, hogy az egyes halakban milyen mutáció okozza a szem elsorvadását, mindenesetre mivel számos vak Astyanax populáció nagyon távol él egymsától, feltételezhető, hogy a párhuzamos konvergens evolúció klasszikus példájával állunk szemben, vagyis a vakság egymástól függetlenül alakult ki. Ennek tesztelésére tökéletesen alkalmas a komplementáció, épp ezért végezte ezt el az NYU egyik kutatója, Richard Borowsky.
Egész pontosan négy különböző barlangi populációt vett górcső alá, ezek a pachóni, a molinoi, a tinajai és a curvai. A komplementáció eredményeképpen a Pachón-Tinaja és Pachón-Molino párosítások funkcionálisan látó (bár a normálisnál kisebb szemű) halakat eredményeztek - nem rossz teljesítmény olyan halpopulációk esetén, ahol több tízezer generáció óta senkinek nem volt működő szeme. Vagyis ezek a halak különböző mutációknak köszönhetik szemtelenségüket. Csak a Curva-Tinaja párosítás nem "működött" - ám mivel ezek a barlangok egyébként földrajzilag közel vannak egymáshoz, elképzelhető, hogy eredetileg az egyik barlangban terjedt el a vak populáció, s majd később, valamilyen földalatti járatokon keresztül jutott át a másik helyre.
Borowsky R (2008) Restoring sight in blind cavefish. Curr Biol 18(1): R23-24.
Niven JE (2008) Evolution: Convergent Eye Losses in Fishy Circumstances. Curr Biol 8(1): R27-R29.