Hogy a nemek milyen arányban oszlanak meg a populáción belül, egyáltalán nem közömbös a közösség jövője szempontjából. Például a szexuálisan szaporodó gerinces fajok esetében az ideális fele-fele aránytól való tartós eltérés, hosszú távon nem a fennmaradást biztosítja, hanem az almanachok "élt, de kihalt" kategóriájába való bekerülést.
Hogy a nemek milyen arányban oszlanak meg a populáción belül, egyáltalán nem közömbös a közösség jövője szempontjából. Például a szexuálisan szaporodó gerinces fajok esetében az ideális fele-fele aránytól való tartós eltérés, hosszú távon nem a fennmaradást biztosítja, hanem az almanachok "élt, de kihalt" kategóriájába való bekerülést.
Nem teljesen véletlen tehát, hogy sok faj nem a szerencsére (értsd: környezeti tényezők) bízza a nemek helyes arányának kialakítását, hanem saját kezébe veszi a sorsát és kialakít egy genetikai alapú nemmeghatározási, azaz szexdeterminációs mechanizmust.
Ennek legkézenfekvőbb példái mi magunk vagyunk: az ún. homogamétás egyedek két, azonos verzióját hordozzák a nemi kromoszómáknak (XX), vagyis nőkké válnak, míg a heterogamétás példányok, a populációban keringő mindkét nemi kromoszómából egyet-egyet mondhatnak magukénak (XY), s így férfiakká válnak. A rendszer kulcsgénje az Y kromoszómán van, ezért ez a kromoszóma élet és halál ura: jelenléte biztosítja, hogy a kezdetleges női nemiszervek sorvadjanak el, férfi ekvivalenseik pedig vidáman tovább fejlődjenek.
Az "XY-rendszer", azonban nem az egyetlen létező, hiszen a madarak és hüllők többsége a fordított logikájú "ZW" mellett tette le a voksát, ahol a homogamétás (ZZ) egyedekből hímek, a heterogamétásokból (ZW) pedig nőstények keletkeznek. (Az eredmény persze ugyanúgy 50-50% nem-eloszlás lesz.)
A fent említett csoportokban, a nemmeghatárózó mechanizmusok nem tűnnek képlékenyek, és általában vagy az "XY-" (emlősök), vagy a "ZW rendszer" (madarak) jellemző az odatartozó összes fajra. Ezzel szemben a kétéltűek közt egyaránt felfedezhetjük mindkét szisztéma képviselőit, sőt olyan faj is akad, ahol közeli populációk (!) különböző módon döntik el, hogy kiből legyen legény, kiből pedig leány.
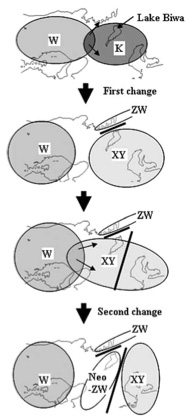 Ide tartozik a Rana rugosa nevű békafaj, amely a Japán-tenger partjain fekvő országokban őshonos. Az már korábban is tisztázott volt, hogy Japánon belül a béka különböző populációi különböző logika szerint határozzák el a nemiséget, de az csak most vált nyilvánvalóvá, hogy pl. a ZW rendszer kétszer, egymástól függetlenül is kialakult.
Ide tartozik a Rana rugosa nevű békafaj, amely a Japán-tenger partjain fekvő országokban őshonos. Az már korábban is tisztázott volt, hogy Japánon belül a béka különböző populációi különböző logika szerint határozzák el a nemiséget, de az csak most vált nyilvánvalóvá, hogy pl. a ZW rendszer kétszer, egymástól függetlenül is kialakult.
Felthetően a R. rugosa legrégebbi populációi, a Nyugat-Japánban (W) ill, a Tokió környéki Kanto régióban (K) őshonosak. Mindkét társaság egy XY alapú rendszert használ, bár ez esetben az X és Y kromoszómák alakjukban nem különböznek egymástól (ellentétben az emberi X-el és Y-al). A két rendszer azonban nem teljesen azonos, ez onnan is tudható, hogy a W békákat keresztezve a K békákkal, torzult nemi arányok jönnek létre, ami, mint fent már említettem nem éppen szép reményekkel kecsegtet. Ezért aztán a két (ősi) populáció átfedésénél keletkező hibridek kénytelenek voltak valamit kitalálni, hogy a génjeik ne menjenek veszendőbe. Az eredmény egyszerre két új rendszer egy XY és egy ZW kialakulása volt.
A történet nem ér még itt véget, ugyanis az újonnan kialakult "XY" populáció egyedei igen promiszkuisnak bizonyultak és páran keveredtek az ősi "W" társaság tagjaival. Az eredmény ismét eltolódott nemi arány lett (egy W és XY béka nászából születő utódok több mint 60%-a hím egy éves korukban), így a rendszer újfenti tuningolásra szorult. Ennek eredményeképpen egy új, az elsőtől különböző "ZW-rendszer" alakult ki az ún. "neo-ZW" populációban. (Hogy a rendszer ennyire képlékeny és XY logikából viszonylag gyorsan lehet ZW logikát fabrikálni, szerintem mindenkit váratlanul ért.)
Az igazán érdekes persze az lesz majd, ha tudni fogjuk, molekulárisan mit is rejtenek a különböző logikájú rendszerek, de erre egy ideig még sajnos várni kell.
Ogata M, Hasegawa Y, Ohtani, Mineyama M, Miura I (2007) The ZZ/ZW sex-determining mechanism originated twice and independently during evolution of the frog, Rana rugosa. Heredity doi: 10.1038/sj.hdy.6801068