A világot nappal (fizikailag) színesnek látjuk, kedvtől függetlenül. Ez, ha felidézzük a korabeli biológia órákat, a retinában található csapsejteknek köszönhető, az pedig, hogy éjszaka is látunk (bár leginkább csak egy színben) az ugyanott levő pálcikasejtek javára írandó.
A világot nappal (fizikailag) színesnek látjuk, kedvtől függetlenül. Ez, ha felidézzük a korabeli biológia órákat, a retinában található csapsejteknek köszönhető, az pedig, hogy éjszaka is látunk (bár leginkább csak egy színben) az ugyanott levő pálcikasejtek javára írandó.
Mindkét sejttípusban (pontosabban mind az ötnégyben, hiszen csapsejtből, attól függően, hogy milyen színt érzékel, három is akad) a fénysugarak érzékelésében kulcsszerepe van az opszin molekuláknak, amelyek sejttípusokként kicsit különböznek egymástól.
Ezek az apró különbségek teszik lehetővé, hogy az egyes retina-sejttípusok opszinjai (és ezáltal maguk a sejtek is), más-más hullámhosszú (vagyis színű) fényre "gerjedjenek". S a csapsejtek három opszinja azért kiemelendő, mert ha csak egyik is hiányozna, nem beszélhetnénk ún. trikromatikus színlátásról egyáltalán.
 Mint ahogy a legtöbb emlős nem is beszélhet, hiszen ebből a szempontból az óvilági majmok (köztük saját közelebbi csoportunk, a főemlősök is) különlegesek a maguk három plusz egy opszinjával (a plusz egy, az a pálcikákban található rodopszin). A többi, "mezei" emlősnek be kell érnie kettő plusz eggyel: a hosszabb hullámhosszú, vörös fényt érzékelő LWS (Long Wavelength Sensitive) opszinnal, a rövid hullámhosszú, kék fényt érzékelő SWS1 (Short Wavelength Sensitive - 1) opszinnal, illetve a már említett rodopszinnal.
Mint ahogy a legtöbb emlős nem is beszélhet, hiszen ebből a szempontból az óvilági majmok (köztük saját közelebbi csoportunk, a főemlősök is) különlegesek a maguk három plusz egy opszinjával (a plusz egy, az a pálcikákban található rodopszin). A többi, "mezei" emlősnek be kell érnie kettő plusz eggyel: a hosszabb hullámhosszú, vörös fényt érzékelő LWS (Long Wavelength Sensitive) opszinnal, a rövid hullámhosszú, kék fényt érzékelő SWS1 (Short Wavelength Sensitive - 1) opszinnal, illetve a már említett rodopszinnal.
A mi extra opszinunk a MWS (Middle Wavelength Sensitive) opszin, amely a zöld szín érzékelésében jeleskedik. (A színtévesztők esetében az LWS vagy MWS opszin szekvenciájában van valamilyen változás, amiért az érzékelési tartományuk elcsúszik, ill. extrém esetelben az egyik opszin teljesen hiányzik - lásd pl. John Dalton. És mivel a két gén az X kromoszómán van, a mutáció sokkal inkább érinti a férfiakat, mint a nőket.) Ezzel azonban tényleg csak a szűkebb "rokonságban" dicsekedhetünk, hiszen jóformán minden más gerincesnek több opszinja van.
Olyannyira, hogy a genom-összehasonlítgatások alapján, ma úgy tűnik, hogy a gerincesek közös őse maga is jó színlátó volt, négy különböző opszinnal. Az LWS és SWS1 mellett SWS2 és Rh2 opszinjuk is lehetett, előbbi a világoskék, míg utóbbi a zöld szín érzékelését teszi lehetővé. Érdekes módon a hipotetikus ősből éppen a szürkületi látásban fontos rodopszin (Rh1) hiányzik, amely egy kicsit később, az állkapcsos gerincesek megjelenésekor jött létre az Rh2 opszin duplikációja során (lásd bal oldali ill. alábbi ábra).
A minimum négy plusz egy opszin leosztás aztán elég standard maradt a legtöbb gerinces csoportban az évmilliók során (néhány halfajban a genomduplikációknak köszönhetően hétre is felment a számuk), egészen az emlősök kialakulásáig, amikor mind az Rh2-, mind az SWS2 opszin génje elveszett - feltehetőleg az emlős-ős éjszakai életével okolható ez, hiszen ezeknek a fehérjéknek így semmilyen szerepük nem maradt. (Az éjszakai életmódhoz kötődő opszin-vesztés mai utódjaikban sem ismeretlen.) A színes gyümölcsökkel táplálkozó óvilági majmokban aztán az LWS opszin gén duplikációjával - ill. az azt követő molekuláris finomhangolással - alakult ki a MWS opszin.
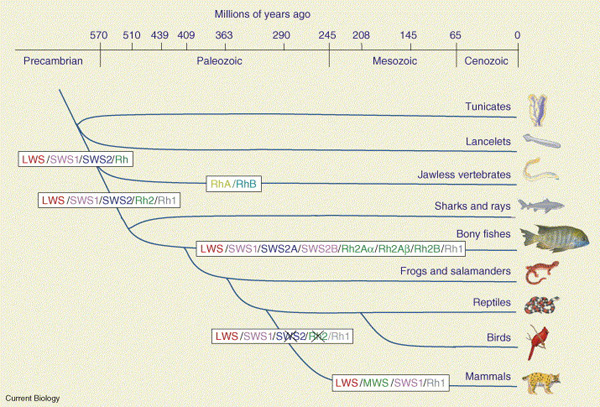
Na, akkor most ezen a fenti képen árnyalunk egy kicsit. Ugyanis a ma élő, legősibb emlőscsoport, a kloakások egyik jeles képviselőjét, a kacsacsőrű emlőst megvizsgálva az derült ki, hogy ő bizony rendelkezik SWS2 opszinnal. (Mondjuk, amit nyer a réven, azt elveszti a vámon, hiszen az SWS1 opszint kódoló génje viszont működésképtelen.) Ez pedig azt jelzi, hogy a mai emlősök közös őséből még csak az Rh2 opszin hiányzott, az SWS1 ill. SWS2 opszin csak a nem kloakás - kloakás emlősök szétválása után veszett el. (Érdekes lesz majd, ha kiderül, hogy az SWS1 hiánya általános kloakás vonás, vagy csak a kacsacsőrű emlős extravaganciája.)
A maradék opszinok aztán a különböző életstílusoknak megfelelő körülményekhez alkalmazkodtak minden gerinces csoportban. Például a tengerek mélyén (is) kódorgó bálnák és delfinek SWS1 opszinja a mélybe lejutó rövidebb fényhullámokra érzékeny, vagy a Viktória-tó zavaros parti vizeiben élő sügérek LWS opszinja a jobban érzékelhető hosszabb hullámhosszok érzékelésére adaptálódott.
Davies, WL, Carvalho, LS, Cowing, JA, Beazley, LD, Hunt, DM, Arrese, CA (2007) Visual pigments of the platypus: A novel route to mammalian color vision. Curr Biol 17: R161-R163.
Trezise, AEO, Collin, SP (2005) Opsins: Evolution in Waiting. Curr Biol 15: R794-R796.