 A biológia különösen érdekes fejezetei közé tartoznak az élősködők és gazdaszervezeteik között zajló millió éves fegyverkezési versenyek. Az élőlények ugyanis az evolúció során a környezetükhöz alkalmazkodnak, azonban ez esetben a környezetük nagyon fontos része egy élőlény is, amely szintén evolvál, így a gazda evolúcióját az élősködője hajtja, az élősködőét meg a gazdáé. Egy ilyen rendszerben sok evolúciós újdonság önmagában előnyt jelent, hiszen minden olyan húzás, ami a gazda/élősködő immunrendszerét készületlenül éri, javíthatja a tulajdonosa túlélési esélyeit. Az egyik szokásos ilyen gazda-élősködő rendszer, amiről viszonylag sok adattal is rendelkezünk a Drosophila melanogaster ecetmuslica és egy élősködő darázs, amely a Leptopilina boulardi névre hallgat. Mindkettő könnyen kezelhető, az ecetmuslica genetikai modellszervezet, az L. boulardi pedig egy egyszerű cukros tápon eltartható, ha néhány nőstényt ecetmuslica lárvákat tartalmazó üvegbe teszünk, szépen megkeresik a lárvákat és megszúrják őket, majd a bejuttatott petékből kikelő darázslárvák belülről felfalják a muslica lárvát. Nyilván ez a folyamat nem száz százalékos hatékonyságú, időnként a darázslárva pusztítja el a muslica lárvát, ekkor a bábozódás után egy darázs kel majd ki, de az is gyakran megtörténik, hogy a muslica lárva immunrendszere győzi le az élősködőt, egy fekete, melanizált tokot képez körülötte, amiben a darázs lárva elpusztul.
A biológia különösen érdekes fejezetei közé tartoznak az élősködők és gazdaszervezeteik között zajló millió éves fegyverkezési versenyek. Az élőlények ugyanis az evolúció során a környezetükhöz alkalmazkodnak, azonban ez esetben a környezetük nagyon fontos része egy élőlény is, amely szintén evolvál, így a gazda evolúcióját az élősködője hajtja, az élősködőét meg a gazdáé. Egy ilyen rendszerben sok evolúciós újdonság önmagában előnyt jelent, hiszen minden olyan húzás, ami a gazda/élősködő immunrendszerét készületlenül éri, javíthatja a tulajdonosa túlélési esélyeit. Az egyik szokásos ilyen gazda-élősködő rendszer, amiről viszonylag sok adattal is rendelkezünk a Drosophila melanogaster ecetmuslica és egy élősködő darázs, amely a Leptopilina boulardi névre hallgat. Mindkettő könnyen kezelhető, az ecetmuslica genetikai modellszervezet, az L. boulardi pedig egy egyszerű cukros tápon eltartható, ha néhány nőstényt ecetmuslica lárvákat tartalmazó üvegbe teszünk, szépen megkeresik a lárvákat és megszúrják őket, majd a bejuttatott petékből kikelő darázslárvák belülről felfalják a muslica lárvát. Nyilván ez a folyamat nem száz százalékos hatékonyságú, időnként a darázslárva pusztítja el a muslica lárvát, ekkor a bábozódás után egy darázs kel majd ki, de az is gyakran megtörténik, hogy a muslica lárva immunrendszere győzi le az élősködőt, egy fekete, melanizált tokot képez körülötte, amiben a darázs lárva elpusztul.
Nem meglepő módon a darázs több különböző módon próbálja gátolni a muslica immunválaszát, a petével együtt különböző faktorokat juttat a muslica lárva testébe, ezek egyikéről lesz most szó, a vírusszerű részecskékről (angolul virus like particle, vagy röviden VLP).
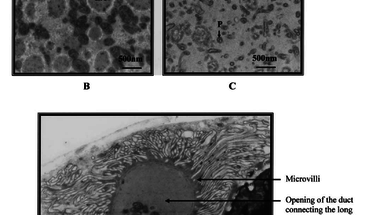
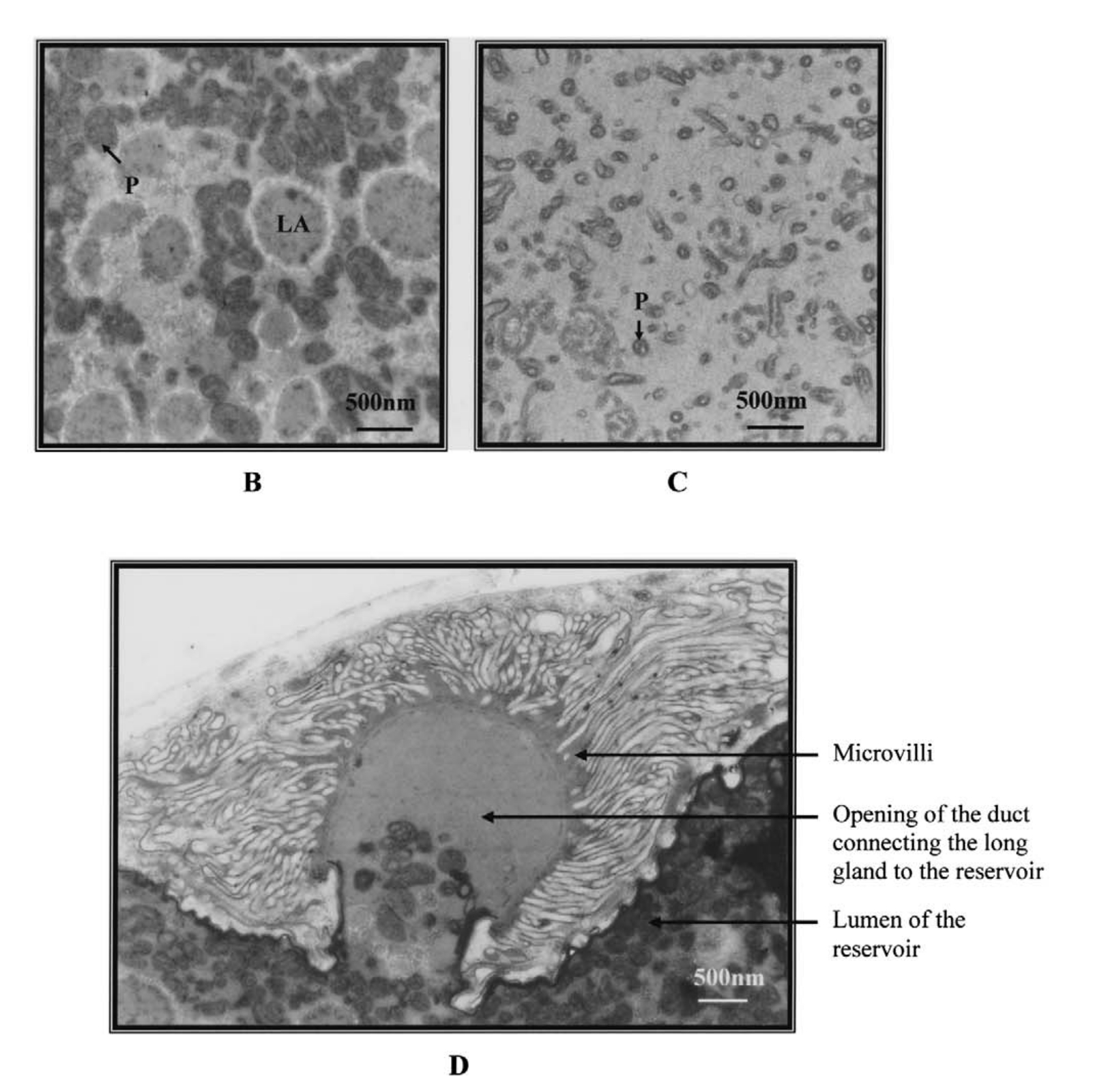
Már a kilencvenes években megfigyelték, hogy a nőstény L. boulardi darazsak hosszú mirigyének (long gland) elektronmiszkroszkópos képein vírusokra hasonlító részecskék figyelhetőek meg, és ezek külalakja korrelál a darázs fertőzőképességével. Ugyanis egy Brazzaville -ből származó L. boulardi törzsről megfigyelték, hogy más külalakú részecskéket tartalmaz és nagyon nehezen képes fertőzni a D. melanogaster lárvákat (ezek láthatóak az első képen, a "P" jelöli őket, a "b" ábrán láthatóak a virulens törzs VLP -i, a "c" ábra pedig a nem-virulens törzs VLP-it mutatja). Egy jó darabig nem is jutottak ennél sokkal tovább.
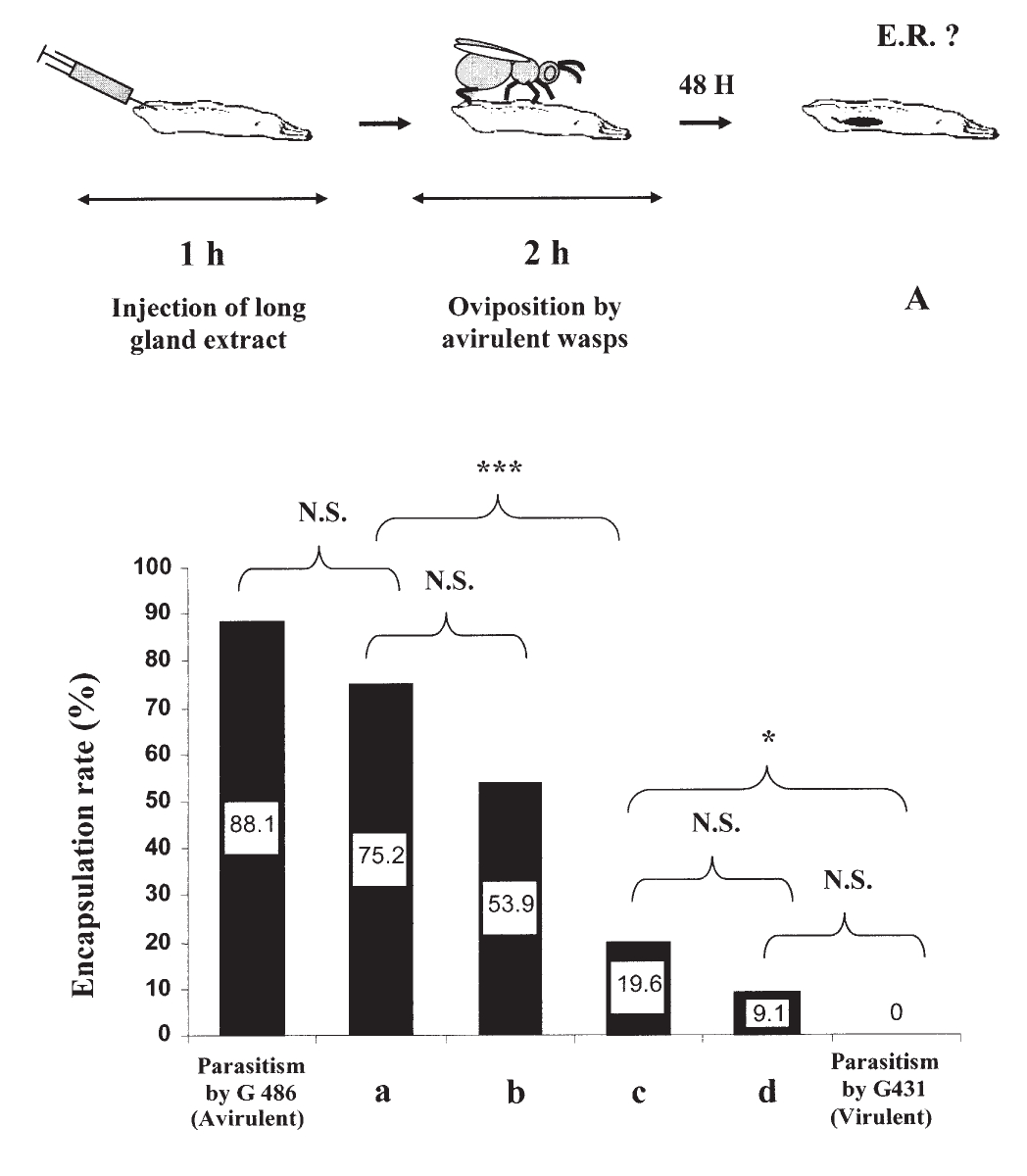
Ám 2003 -ban Labrosse és munkatársai egy közleményben pofonegyszerű módon vizsgálták ezt a folyamatot. Ha a hatékonyan fertőző, virulens darázstörzset petéztették muslica lárvákba, a darázspeték körül nem alakult ki a fekete, melanizált tok. Ellenben a rossz hatékonysággal fertőző, nem-virulens darázstörzs petéinek nyolcvan százaléka körül kialakult a fekete, melanizált tok. Eddig ebben semmi meglepő sincs, viszont ha a muslica lárvákat először a virulens darázzsal szúratták meg, majd utána a nem-virulenssel, akkor azt tapasztalták, hogy a petéknek csak öt százaléka feketedik meg, tehát nyilvánvalóan a virulens darázs valamilyen módon gátolja a gazda immunválaszát, amire a nem-virulens darázs nem képes. Ezek után fogták a virulens darazsakat és a kibontott hosszú mirigyeik váladékát muslica lárvákba oltották, majd ez után hagyták, hogy a nem-virulens törzs nőstényei petéket rakjanak beléjük. Ez látható a második ábrán, az első oszlopban a nem-virulens darazsak petéztek az ecetmuslica lárváiba. Az "a" oszlopban a nem-virulens darazsak mirigyváladékát oltották a muslica lárvákba, majd a nem-virulens darazsak petéztek beléjük, látható, hogy a darázspeték nagy része megfeketedik, a muslica az esetek többségében sikeres immunválaszt adott. A "b" oszlopban látható kísérletben a nem-virulens darázstörzs egyedei petéztek a muslica lárvákba, majd ez után oltották be őket a virulens törzs mirigyváladékával, ez láthatóan növeli a darázsfertőzés hatékonyságát. A "c" kísérlet ugyanez, csak előbb oltották be a mirigyváladékot és utána petéztettek. A "d" oszlopban ugyanez a kísérlet látható, de a mirigyváladékot beoltás előtt fél órán át hetven fokra hevítették, láthatóan ez nem csökkenti a hatékonyságát. Ezekből a kísérletekből nyilvánvaló, hogy a hosszú mirigy váladéka akadályozza valahogyan a gazda immunválaszát.
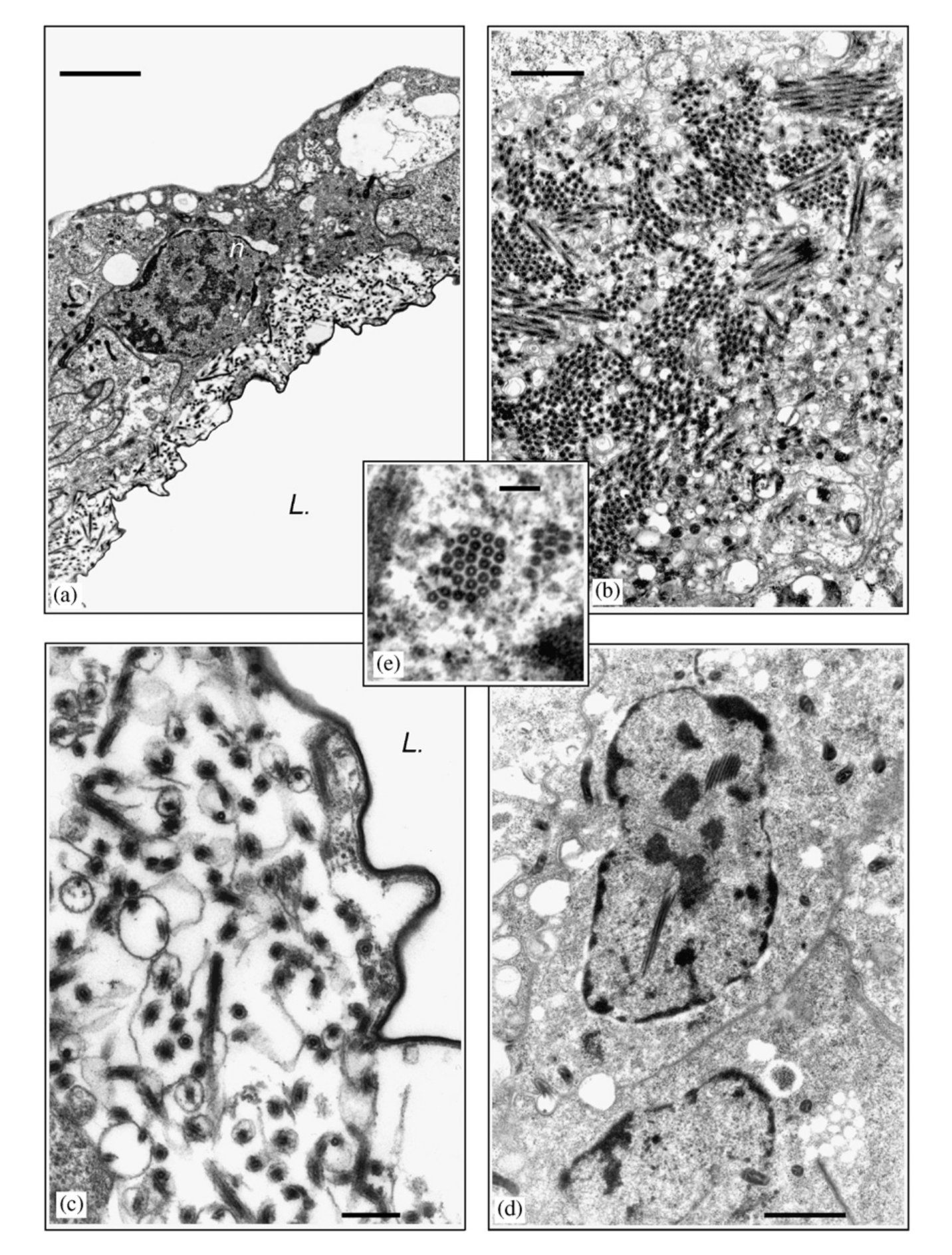
Varaldi és munkatársai 2006 -ban hasonló kísérletekkel ugyanerre a következtetésre jutottak, miszerint a hosszú mirigy váladéka felelős a gazda immunrendszerének gátlásáért. Viszont elektronmikroszkóppal jó alaposan végignézték a nőstény darazsak petefészkeit, a petefészek üregét bélelő sejtekben találtak is a sajtmagokban szaporodó vírusokat, innen feltételezték, hogy egy DNS vírusról lehet szó. Ezek láthatóak a harmadik képen.
Martinez és munkatársai 2011 -ben közölték le azt az eredményt, ami az egész történetnek még egy csavart ad. Ugyanis ennek a szimbionta vírusnak a felfedezésével egy újabb elemmel bővült a rendszer, nyilván az ecetmuslica, mint gazda és a darázs, mint élősködő mellett a darázs szimbionta vírusai is a többi kettőtől függetlenül evolválnak. Úgyhogy a szerzők kimentek terepre és Franciaország déli részeiről begyűjtöttek egy csomó L. boulardi törzset, amelyek közt volt a VLP vírussal fertőzött és attól mentes is. Ezek után megvizsgálták, hogy ezek miben különböznek egymástól. Meglepő módon azt találták, hogy a vírusfertőzött darazsak gyakran egynél több petét is tojnak egy muslica lárvába, olyan törzs is akadt, amely átlagosan tizenegynél is több petét rakott egyetlen muslica lárvába. Ez a darázs számára nem túl előnyös, mivel egyetlen muslicában csak egyetlen darázs marad életben, ha több darázs lárva kerül egy gazdába, azzal kezdik, hogy elpusztítják egymást, amíg csak egy marad, így ha tizenegy pete kerül egyetlen gazdaegyedbe, abból tíz teljesen fölösleges. Ez látható a negyedik képen.
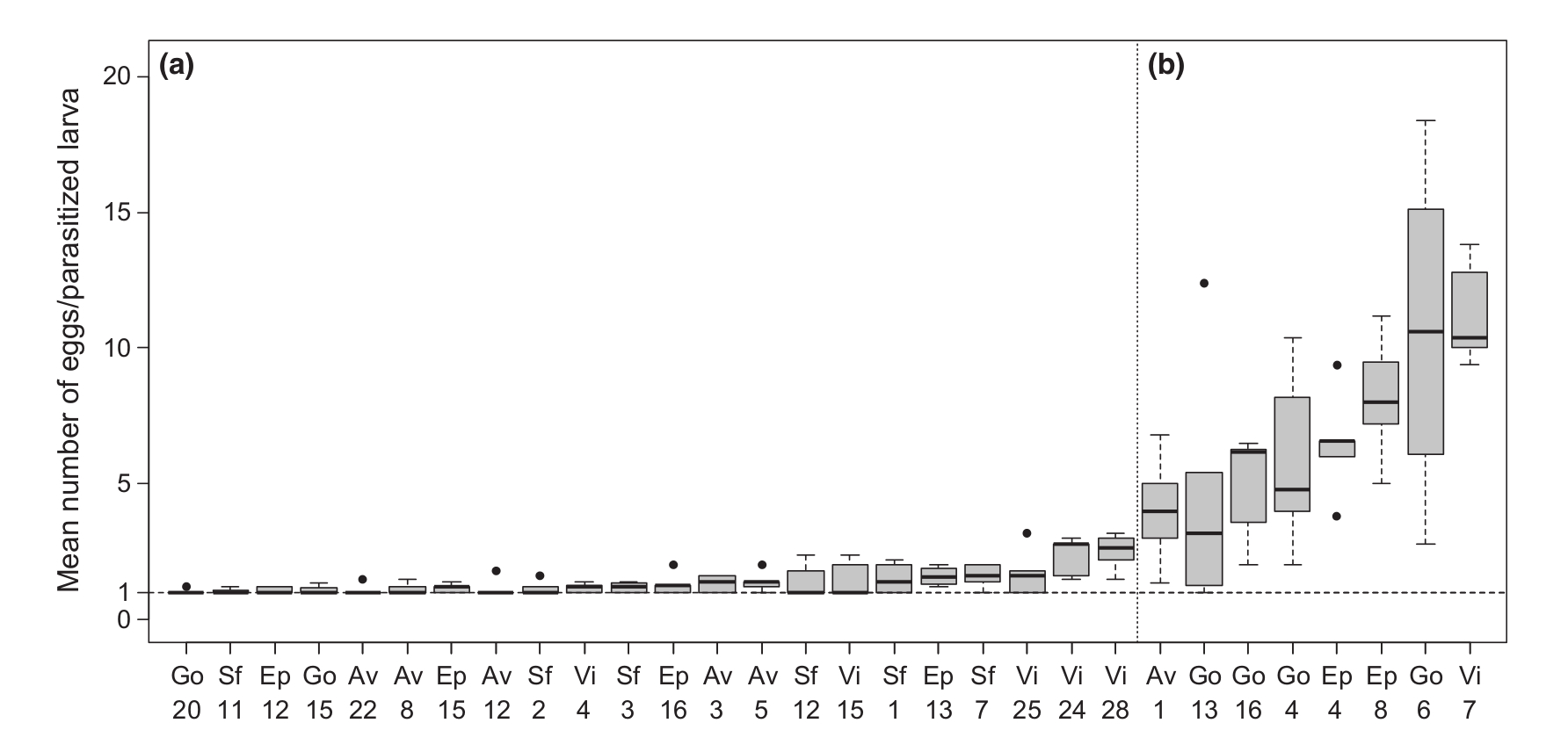
A kérdés az, hogy ez miért előnyös a darázsnak, hiszen egy csomó petét rak le potyára? A darázsnak nem előnyös, azonban a vírusnak igen, a vírus alapvetően egy hordozó nőstény utódait fertőzi, azonban ha egyetlen gazdába két különböző nőstény petéi kerülnek, a vírus átkerülhet az eddig vírusmentes lárvába is, ha az él túl, már ő maga is terjeszteni fogja a szimbionta vírust. Ez nyilván egy evolúciós tradeoff, a vírusfertőzésnek előnyei is vannak (lecsendesíti a gazda immunrendszerét) de hátrányai is jelentkeznek (petepocsékolás), amíg az előnyei meghaladják a hátrányait, a fennmaradására ht szelekciós nyomás.
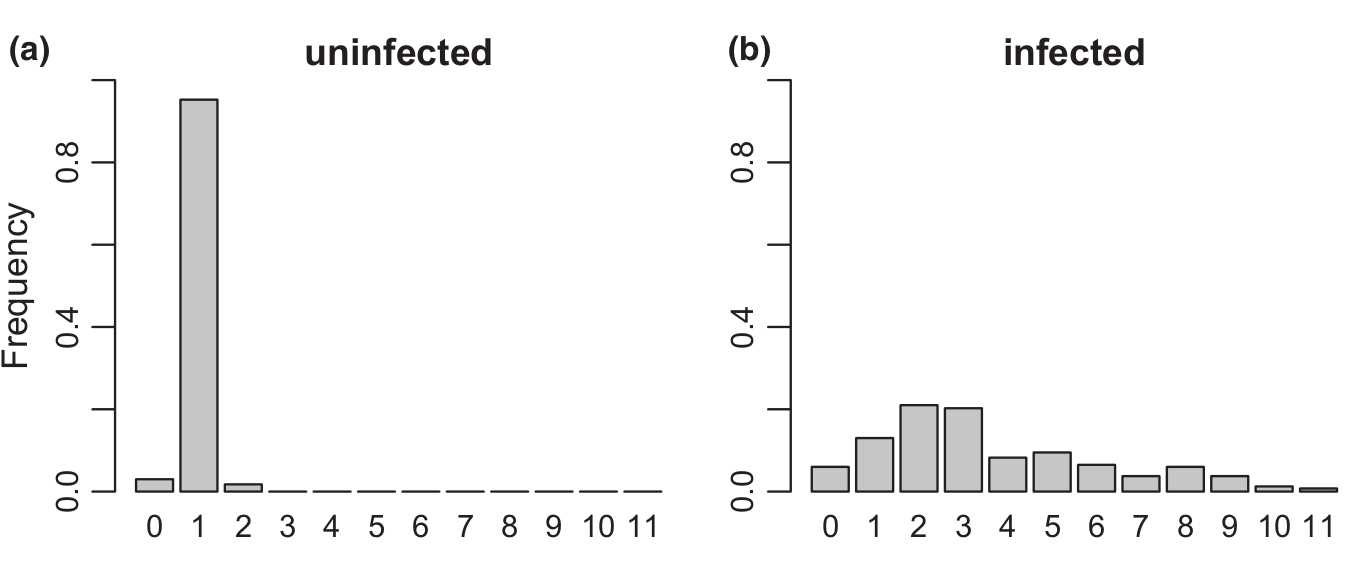
Azonban Patot és munkatárai 2012 -ben újabb csavart vittek ebbe a történetbe: Azt vizsgálták, hogy a vírusfertőzés hogyan befolyásolja az L. boulardi darázs versengését más élősködő darázsfajokkal? Példaként az 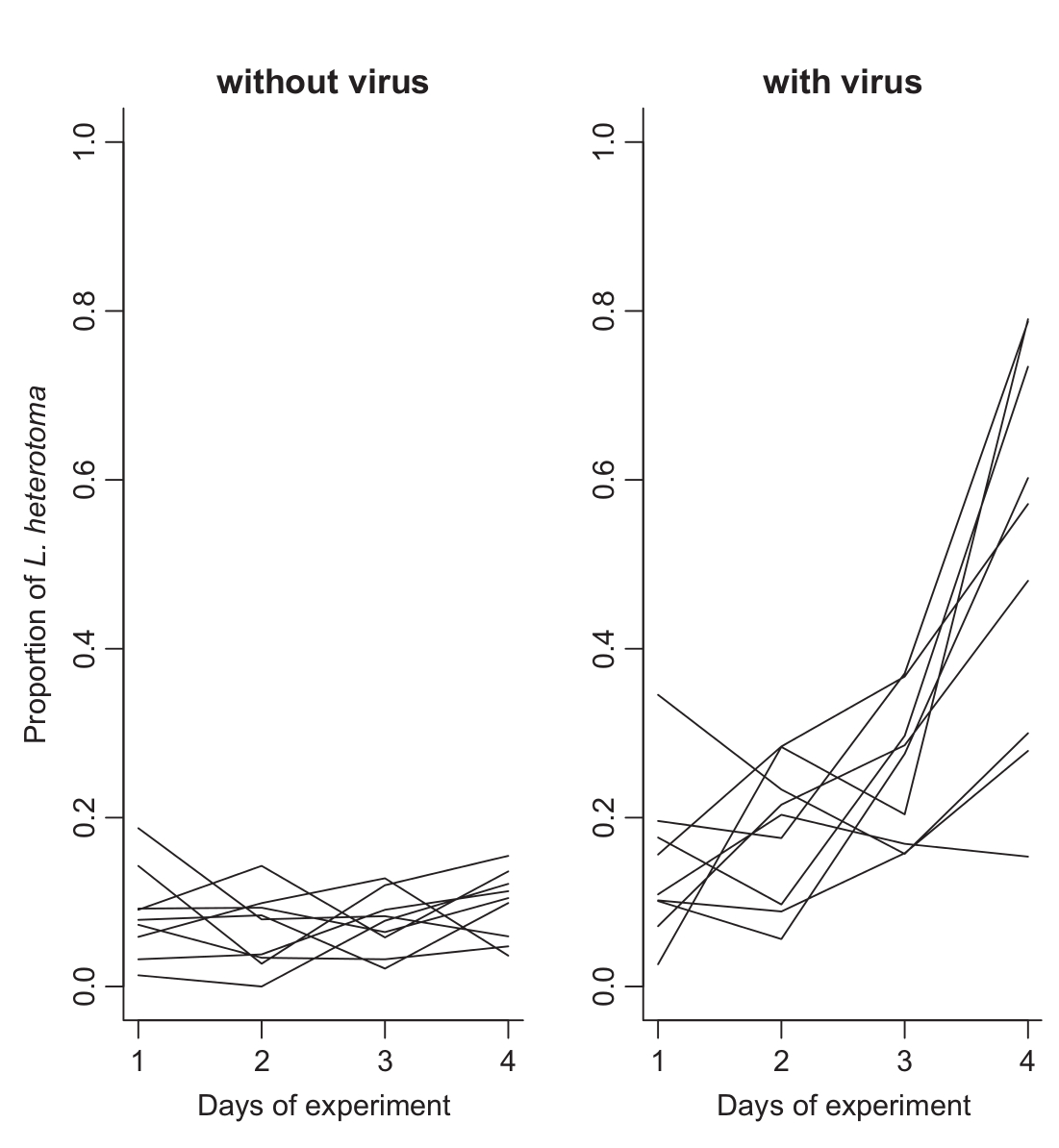 L. heterotoma darázsfajt választották, mert Franciaország déli részén a természetben ez a két leggyakoribb élősködője a D. melanogasternek és gyakran előfordul, hogy egyetlen muslica lárvába mindkét faj belepetézik, így nyilván a természetben is versengenek egymással. Laboratóriumi körülmények között is hajlandóak a másik faj által fertőzött muslica lárvákba petézni, úgyhogy a kísérletek könnyedén elvégezhetőek ellenőrzött körülmények közt is. Azonban az eddig vizsgált vírus, ami az LbFV névre hallgat egyébként (Leptopilia boulardi Filamentous Virus) valamiért nem fertőzi az L. heterotoma fajt, így az L. heterotoma egyedek mindig vírusmentesek. Ők is ugyanazt tapasztalták, mint az előző közlemény szerzői, a vírussal fertőzött L. boulardi nőstények egy muslica lárvába több petét raktak, a vírusmentesek pedig csak egyet, ez látható az ötödik képen.
L. heterotoma darázsfajt választották, mert Franciaország déli részén a természetben ez a két leggyakoribb élősködője a D. melanogasternek és gyakran előfordul, hogy egyetlen muslica lárvába mindkét faj belepetézik, így nyilván a természetben is versengenek egymással. Laboratóriumi körülmények között is hajlandóak a másik faj által fertőzött muslica lárvákba petézni, úgyhogy a kísérletek könnyedén elvégezhetőek ellenőrzött körülmények közt is. Azonban az eddig vizsgált vírus, ami az LbFV névre hallgat egyébként (Leptopilia boulardi Filamentous Virus) valamiért nem fertőzi az L. heterotoma fajt, így az L. heterotoma egyedek mindig vírusmentesek. Ők is ugyanazt tapasztalták, mint az előző közlemény szerzői, a vírussal fertőzött L. boulardi nőstények egy muslica lárvába több petét raktak, a vírusmentesek pedig csak egyet, ez látható az ötödik képen.
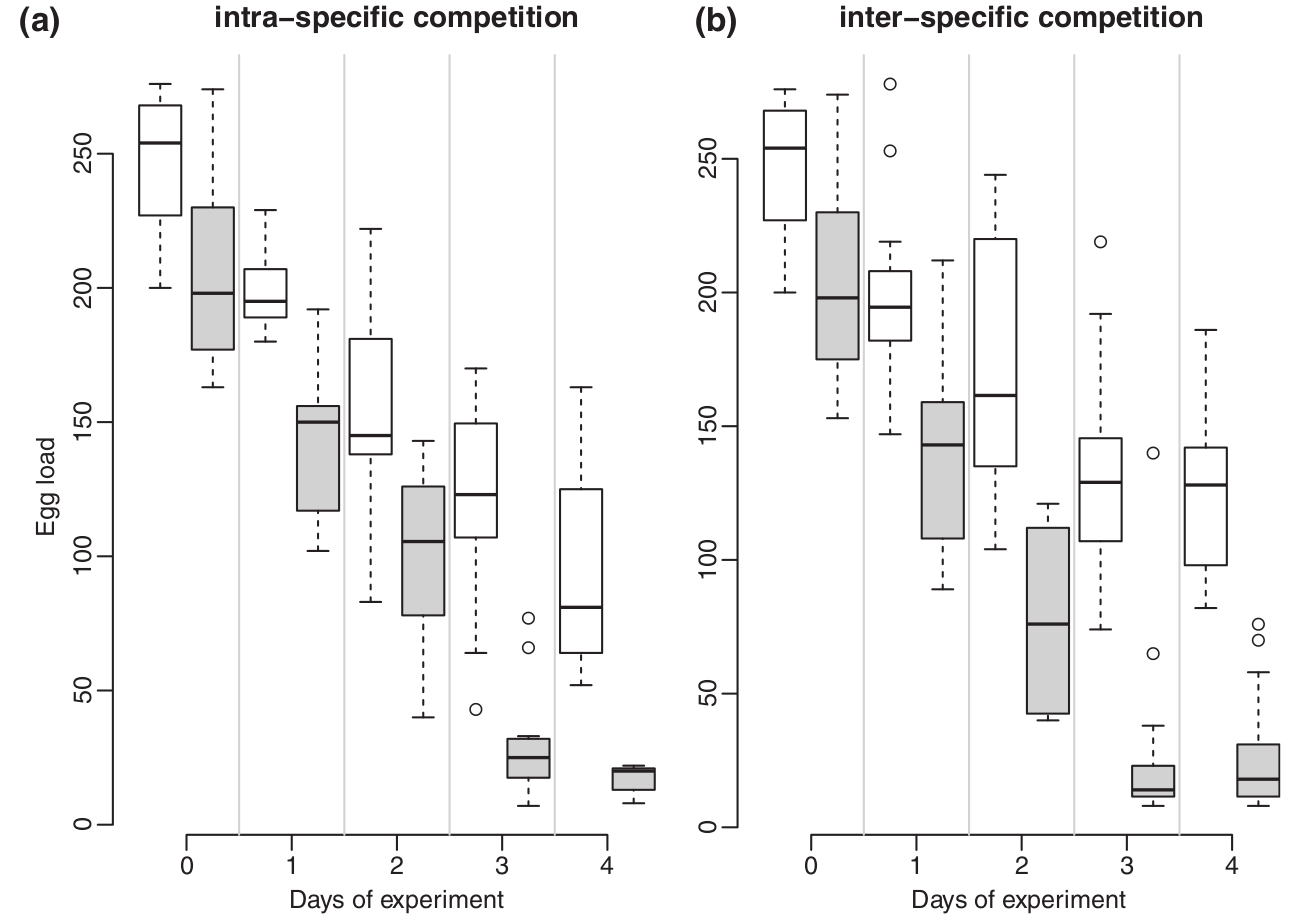
Meglepő módon a vírusmentes L. boulardi erőteljesen kompetált a L. heterotomával, ha négy-négy nőstényt zártak egy üvegbe egy csomó muslica lárvával, a kikelő darazsaknak csak nyolc százaléka bizonyult L. heterotomának. Azonban ha vírusfertőzött L. boulardi nőstényeket választottak, ez az arány megfordult, az L. heterotoma sokkal sikeresebben fertőzte a muslicákat. Ez látható a hatodik ábrán az egyes görbék a kikelő L. heterotoma egyedek arányát mutatják. Meglepő módon azt tapasztalták, hogy a kísérlet során az L. boulardi nőstények (szürke négyszögek) petefészkeiből elfogytak a peték, a negyedik nap végére már jelentősen kevesebb petét tartalmaztak, mint az L. heterotoma nőstények (fehér négyszögek). Ez látható a hetedik ábrán.
Ezek alapján a vírusfertőzött L. boulardi nőstények azért maradnak alul a versengésben, mert egyszerűen elpazarolják a petéiket, amikor egy lárvába többször tojnak, ami mint láttuk egyértelműen a vírusfertőzés következménye. Magyarul, ha az L. boulardinak egy másik darázzsal kell versenyeznie, akkor az LbFV fertőzés minden előnye sem tudja ellensúlyozni a hátrányát, hogy a sok lerakott pete fölösleges pazarlás, így a fertőzött populációk hátrányba kerülnek egy olyan fajjal szemben, amelynek egyébként esélye sem lenne kompetálni velük. Valószínűleg ez a magyarázata, hogy Franciaországban a D. melanogaster mindkét élősködője egyszerre létezik, bár ugyanazt a niche -t foglalják el az LbFV vírus fenntartja a egyensúlyukat. Tökéletes példa ez arra is, hogy a biológiában értelmetlen előnyös vagy hátrányos jellegekről beszélni, rengeteg tulajdonság a környezettől függően előnyös és hátrányos is lehet.
Labrosse C., Carton Y., Dubuffet A., Drezen J.M., Poirie M. (2003): Active suppression of D. melanogaster immune response by long gland products of the parasitic wasp Leptopilina boulardi. Journal of Insect Physiology 49: 513–522
Varaldia J., Ravallecb M., Labrossec C., Lopez-Ferberd M., Boulétreaua M., Fleurya F. (2006) Artifical transfer and morphological description of virus particles associated with superparasitism behaviour in a parasitoid wasp. Journal of Insect Physiology 52: 1202–1212
Martinez J., Fleury F., Varaldi J. (2011) Heritable variation in an extended phenotype: the case of a parasitoid manipulated by a virus. Journal of Evolutionary Biology 25: 54-65
Patot S., Allemand R., Fleury F., Varaldi J. (2012): An inherited virus influences the coexistence of parasitoid species through behaviour manipulation. Ecology Letters 15:603-610