Egy-egy bálnacsontvázzal szembesülve az átlagember első (és gyakran egyetlen) gondolata, hogy "hát igen, ez valóban bazi nagy". Azonban a fizikai nagyság el- és felismerése mellett gyakran hajlamosak vagyunk elsuhanni az apróbb részletek mellett (persze néha kontár preparátorok is segítenek ebben).
Egy-egy bálnacsontvázzal szembesülve az átlagember első (és gyakran egyetlen) gondolata, hogy "hát igen, ez valóban bazi nagy". Azonban a fizikai nagyság el- és felismerése mellett gyakran hajlamosak vagyunk elsuhanni az apróbb részletek mellett (persze néha kontár preparátorok is segítenek ebben).
Éppen egy masszív csontváz kapcsán beszélni "apró" részletekről, furcsának tűnhet (és valóban, minden relatív), de ez esetben mégsem inadekvát. Arról a két kisméretű csontról lesz itt ugyanis (javarészben) szó, amelyek a test két oldalán, a gerincoszlop alatt helyezkednek el a bálna hasában.
A két csont annyira jelentéktelennnek tűnik, hogy kevésbé figyelmes szemlélők gyakran észre sem veszik őket (és nota bene, a bálna mindennapjai szempontjából tényleg nem is bírnak jelentőséggel), pedig evolúciós szempontból egyáltalán nem érdektelenek: arról a korról tanúskodnak, amikor a mai bálnák ősei a szárazföldön jártak. A két csont ugyanis a bálnák medenceövének és hátsó végtagjának a maradéka.
A cetek különösen jól alkalmazkodtak a vízi élethez, hiszen azon kevés emlősfaj közé tartoznak, amelyek képesek egész életüket a vízben leélni: itt táplálkoznak, párzanak, sőt itt hozzák világra utódaikat is. Kevés kétség fér ahhoz is, hogy a bálnák és delfinek testalkata hidrodinamikailag szinte tökéletes, olyan mintha erre az életformára "teremtettek" volna.
Persze "teremtő aktusról" nincs (és nem is volt) szó, de a cetek különlegessége az emlősök között sokáig komoly fejfájást okozott Darwinnak és remek támadó felületet kínált a kreacionista mozgalmaknak az evolúcióval szemben. Az elmúlt évtizedben aztán minden megváltozott. Számos olyan fosszília került napvilágra, amelyek segítségével egyre finomabb képet kaptunk arról, miként is szánta rá magát az emlősök egy csoportja, hogy áttérjen a vízi életre. S bár nyilvánvalóvá vált, hogy Drawin megérzése, miszerint a cetek egy medveszerű ragadozó őstől számaznak pontatlan, mégsem tévedett nagyot. A történet valóban egy ragadozóval kezdődik, egy patás ragadozóval.
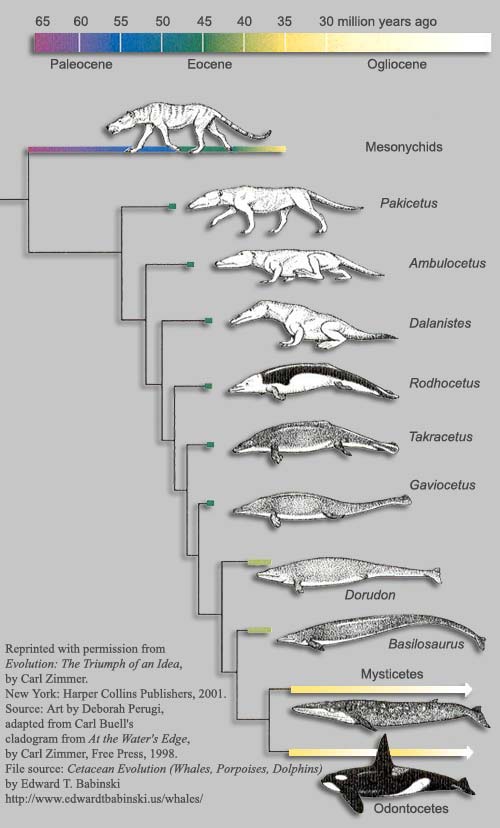 Ez a ragadozó, a mára már kihalt, tartozó Mesonychia csoportba tartozott. Egy farkas alkatú és méretű páros-ujjú patások volt, amelynek jellegzetes fogazata sokban hasonlított a mai ragadozó cetek fogaihoz.
Ez a ragadozó, a mára már kihalt, tartozó Mesonychia csoportba tartozott. Egy farkas alkatú és méretű páros-ujjú patások volt, amelynek jellegzetes fogazata sokban hasonlított a mai ragadozó cetek fogaihoz.
A legősibb szárazföldi "cet" a Pakicetus, amely cirka 50 millió éve örvendeztette meg jelenlétével a környezetét, és a maradványai alapján mindenapi zsákmányát a folyókban halászva szerezte. Őt követi a sorban az Ambulocetus, amely már inkább egy emlősbe oltott krokodilként írható le: állkapcsa megnyúlt és hosszú farka volt, amelyet (feltehetőleg) előszeretettel használt az úszás során.
Ezután ahogy közeledünk korban napjainkhoz, úgy válnak a fosszíliák is egyre delfin- és bálnaszerűbbé. Egyre izomsabb farokúszójuk van, de ugyanakkor még mindegyiken jól felismerhető a hátsó végtag (bár fokozatosan csökken a mérete). Éppen ezért a legtöbb kutató azt valószínűsiti, hogy akárcsak ma a fókák és rozmárok, a Gaviocetus és rokonai még ki-kijöttek a szárazföldre, de ott egyre sutábban mozogtak. A sorozat a Basilosaurusnál látszik megszakadni, ahol már csak egészen elsatnyult hátsó végtaggal találkozunk - ennek már csak a párzásban lehetett szerepe.
(A vízi életmódhoz történő alkalmazkodás során a változás persze nem csak a hátsó végtagra korlátozódott. Jól nyomon követhető a fogazat átalakulása, a légzőnyílás "vándorlása" az állkapocs elejétől a koponya középső részére, vagy éppen a középfül átalakulása. Utóbbi különösen fontos lehetett, hiszen főleg rossz látótávolság esetén a vízben megnő a hallás jelentősége (még echolokáció hijján is), de a "hagyományos" szárazföldi életmódhoz alkalmazkodott emlős hallókészülék felkészületlen a vízben történő tájékozódásra. A külső hallójárat jelentősége elvesztődik, mivel a vízben terjedő hanghullám "berezgeti" az egész koponyát, ez pedig megnehezíti a hang irányának meghatározását. Ezért a cetek hallókészüléke egy párnázott üregben helyezkedik el, ahova az állkapocs körül elhelyezkedő különleges zsírpárna vezeti a hangot.)
 Egy fejlődésbiológus számára persze a legizgalmasabb kérdés, hogy milyen molekuláris és genetikai okokra vezethető vissza a hátsó végtag elvesztése? Mivel sem bálna-, sem delfin embriókkal nem egyszerű dolgozni, a kérdés megválaszolása sokáig váratott magára, de idén tavasszal született egy magyarázat.
Egy fejlődésbiológus számára persze a legizgalmasabb kérdés, hogy milyen molekuláris és genetikai okokra vezethető vissza a hátsó végtag elvesztése? Mivel sem bálna-, sem delfin embriókkal nem egyszerű dolgozni, a kérdés megválaszolása sokáig váratott magára, de idén tavasszal született egy magyarázat.
(Mivel lusta vagyok az alapoktól elindulni ;-), a következő bekezdés tartalmának kisilabizálásához meleg ajánlom a végtagfejlődéses post (újra)olvasását.)
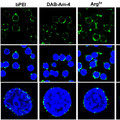
A bálna fosszíliák felkutatásában is jeleskedő Hans Thewissen csoportja pettyes delfinek fejlődő embrióiban kezdte vizsgálni, hogy mely pontjain ment gallyra a végtagfejlődés mechanizmusa.
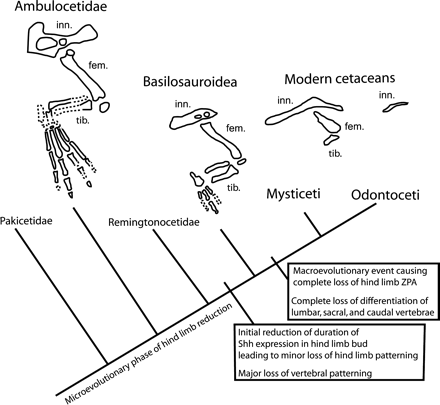
Az már régóta ismert volt, hogy a cetek embrionális fejlődése során megjelennek a hátsó végtagbimbók is, ám ahelyett hogy teljes értékű lábat hoznának létre egy idő után elsorvadnak. Az említett kísérletsorozat során kiderült, hogy ez egyebek mellett azzal magyarázható, hogy a végtagbimbó legfőbb "szervező központjaként" működő apikális ektodermális redő (AER) csak időleges alakul ki, egy idő után eltűnik. További érdekesség, hogy a végtag polaritásának kialkulásában kulcsfontosságú ZPA sem jön létre. Ez utóbbi persze lehetne azzal is magyarázható, hogy amiután az AER már nem alakult ki, a későbbi hátsó végtagfejlődési folyamatokban zavartalanul felhalmozódhattak a mutációk, de ha a fosszíliákat is megnézzük, akkor más következtetésre jutunk.
Sokkal valószínűbb ugyanis, hogy a ZPA hatómolekulájaként működő Sonic-hedgehog koncentrációjának lassú csökkenése okozta a hátsó végtag elsatnyulását az évmilliók során. Majd, amikor a hátsó végtag jelentősége teljesen elvesztődött bekövetkezhetett az AER funkcióvesztése is.
Mint minden evodevos cikkben, itt is fontos hangsúlyozni, hogy a változások/mutációk nem elsősorban a gének fehérje kódoló régióiban történtek, hanem a gének expresszióját szabályozó DNS szekvenciákban. (Mivel ezek a gének rengeteg más fejlődési folyamatban is kiemelten fontosak, ez érthető is.) Ez viszont egyszerűbbé teszi, hogy időnként "visszafordíthatóvá" váljon a változás, és olyan bálnák szülessenek, amelyeknek jól látható lába van. Az ilyen rég "elfelejtett" fejlődési mechanizmusok újraéledése vezet az atavizmusok kialakulásához.
Bővebben:
Talk.Origins: The Origin of Whales and the Power of Independent Evidence
Thewissen labor | Edward Babinski honlapja
Carl Zimmer: At the Water's Edge
Thewissen JG, Cohn MJ, Stevens LS, Bajpai S, Heyning J, Horton WE Jr. (2006) Developmental basis for hind-limb loss in dolphins and origin of the cetacean bodyplan. PNAS 103(22): 8414-8418.
Nummela S, Thewissen JGM, Bajpal S, Hussain ST, Kumar K (2004) Eocene evolution of whale hearing. Nature 430: 776-778.
Thewissen, J. G. M., Bajpai S. (2002) Whale origins as posterchild for macroevolution. BioScience 5: 1037-1049.
Előzmények:
„Kígyónak lábsó...” - Csökevény szervek 1. | Kilóg a kígyóláb
"... madaraknak fogsor" - Csökevény szervek 2. | Re: "...madaraknak fogsor"