A genetikailag módosított szervezetek (GMO-k) kapcsán való álláspontunknak, ill. a Greenpeace-el való egyetnermértésünket már többször hangot adtam, aminek egyeneságú következménye, hogy a szóban forgó szervezet elkötelezett és kritikát nem tűrő rajongóinak szemében erkölcstelen GMO-lobbisták lettünk.
A genetikailag módosított szervezetek (GMO-k) kapcsán való álláspontunknak, ill. a Greenpeace-el való egyetnermértésünket már többször hangot adtam, aminek egyeneságú következménye, hogy a szóban forgó szervezet elkötelezett és kritikát nem tűrő rajongóinak szemében erkölcstelen GMO-lobbisták lettünk.
Igaz, mindössze annyit állítottunk (és állítunk ma is), hogy:
"nincs semmilyen a priori okunk arra, hogy a génmódosítást félelmetesnek állítsuk be, és a génmódosítás, mint technológia, csuklóból elutasítása nemcsak alaptalan és tudománytalan, de felelőtlen is. [...] A zöld mozgalmak elemi érdeke, hogy egy flottul működő teszt- és monitoring rendszert állítsanak fel, amivel kiszámíthatóan lehet tesztelni az egyes GM fajokat, és ezek alapján a valóban veszélyeseket betiltani, a többit meg szépen engedni termeszteni. Ha "erkölcsi alapon" született dogmatikus döntésekhez akarjuk igazítani a tudományos eredményeket, akkor azzal magunkat és a tudományos módszert járatjuk le."
Ez viszont épp elég vörös posztó azoknak a kvázi ludditáknak, akik valamiért hiszik és vallják, hogy a gén módosított növény (vagy állat) az csak szörnyű lehet. Nehéz ennek a miértjét felfogni, hiszen az egyetlen magyarázat az lehetne, hogy valamiféle genom-szintű sztázisban hisznek, abban, hogy az élő szervezetek genetikai anyaga nem változik folyamatosan. Ez természetesen abszurd, és ha jobban megnézzük, nagyon sok elképesztően veszélyesnek és felelőtlennek beállított manipulációnak megtaláljuk a természetes megfelelőjét.
A genetikailag módosított szervezetek (GMO-k) kapcsán való álláspontunknak, ill. a Greenpeace-el való egyetnermértésünket már többször hangot adtam, aminek egyeneságú következménye, hogy a szóban forgó szervezet elkötelezett és kritikát nem tűrő rajongóinak szemében erkölcstelen GMO-lobbisták lettünk.
Igaz, mindössze annyit állítottunk (és állítunk ma is), hogy:
"nincs semmilyen a priori okunk arra, hogy a génmódosítást félelmetesnek állítsuk be, és a génmódosítás, mint technológia, csuklóból elutasítása nemcsak alaptalan és tudománytalan, de felelőtlen is. [...] A zöld mozgalmak elemi érdeke, hogy egy flottul működő teszt- és monitoring rendszert állítsanak fel, amivel kiszámíthatóan lehet tesztelni az egyes GM fajokat, és ezek alapján a valóban veszélyeseket betiltani, a többit meg szépen engedni termeszteni. Ha "erkölcsi alapon" született dogmatikus döntésekhez akarjuk igazítani a tudományos eredményeket, akkor azzal magunkat és a tudományos módszert járatjuk le."
Ez viszont épp elég vörös posztó azoknak a kvázi ludditáknak, akik valamiért hiszik és vallják, hogy a gén módosított növény (vagy állat) az csak szörnyű lehet. Nehéz ennek a miértjét felfogni, hiszen az egyetlen magyarázat az lehetne, hogy valamiféle genom-szintű sztázisban hisznek, abban, hogy az élő szervezetek genetikai anyaga nem változik folyamatosan. Ez természetesen abszurd, és ha jobban megnézzük, nagyon sok elképesztően veszélyesnek és felelőtlennek beállított manipulációnak megtaláljuk a természetes megfelelőjét.
Hely és idő hiányában nem vesézném végig egyenként az összes hibás vagy értelmezhetetlen érvet egyenként, amit a Greenpeace előzékenyen felsorol a "GM: kérdések és válaszok" honlapján, inkább az összhatásra koncentrálnék. Az érvük, egy mondatban, valahogy így szólna: a génmódosítás során bejutó "idegen" DNS darab integrációja véletlenszerű és kiszámíthatatlan hatásai vannak, s így a megváltozott genomú növény veszélyes lesz.
Mindez logikusnak hangzik, amíg nem vagyunk tisztában az eukarióta (így növényi) genomok dinamikájával. Ugyanis a kisebb-nagyobb kromoszomális átrendeződések, transzpozon inszerciók folyamatosak, így állandóan változó, egymástól különböző genomok jelennek meg a természetben is. Valahogy mégsem akarunk minden egyes paradicsomtő mellé ellenőrt rakni, hogy leellenőrizzük, adott génkombináció nem veszélyes-e.
Jó, jó, de hát a GP kifejezetten "idegen" DNS darabokról beszél, az mégsem mindennapos, lehetne ellenvéleményként felhozni. De, viszonylag az. És itt nem is arra gondolok, hogy távoli rokon növények képesek a természetben DNS darabokat átadni, vagy amikor növényi oltás során egymással nem szaporodó fajok cserélnek DNS darabokat az oltási felületen (s lám mégsem hallatszik velük kapcsolatban a "tilcsákbea" kórus...). Hanem prokarióta eredetű DNS szekvenciák folyamatos, ellenőrizhetetlen beilleszkedésére a növényi genetikai anyagba. Ami, ha jól értelmezem mondanivalójukat, a GP aktivisták szerint a lehető legveszélyesebb dolog.
![]() Az talán senki számára nem sokkoló újdonság, hogy a növényi sejtben a sejtmagon kívül is van DNS, egész pontosan a mitokondriumokban illetve a kloroplasztiszokban. Ez a két sejtorganellum még az evolúció korai fázisában lépett szimbiózisra a (proto)eukarióta sejttel, és mára már annyira gördülékeny lett ez az együttélés, hogy saját (jellegükben prokarióta) génjeik nagy része "átköltözött" a sejtmagba (ez dióhéjban az endoszimbiózis elmélete). Mindez persze nem triviális változás, hiszen az átkerülő prokarióta génnek egy megfelelő eukarióta promotert kellett "beszereznie", olyan szekvenciadarabokat "felszednie", amelyek lehetővé teszik, hogy később visszajusson az organellumba, és nem utolsósorban mindezenközben működőképesnek maradnia. Minden valószínűség szerint nem is sikerült általában elsőre az "átugrás", temérdek sikertelen kísérlet kellett minden egyes sikeres endoszimbiotikus géntranszferhez. Ami viszont fel is veti a kérdést, hogy a sikertelen alkalmakat is beleszámítva, mennyire gyakori a plasztisz, ill. mitokondriális eredetű gének átjutása a sejtmagba?
Az talán senki számára nem sokkoló újdonság, hogy a növényi sejtben a sejtmagon kívül is van DNS, egész pontosan a mitokondriumokban illetve a kloroplasztiszokban. Ez a két sejtorganellum még az evolúció korai fázisában lépett szimbiózisra a (proto)eukarióta sejttel, és mára már annyira gördülékeny lett ez az együttélés, hogy saját (jellegükben prokarióta) génjeik nagy része "átköltözött" a sejtmagba (ez dióhéjban az endoszimbiózis elmélete). Mindez persze nem triviális változás, hiszen az átkerülő prokarióta génnek egy megfelelő eukarióta promotert kellett "beszereznie", olyan szekvenciadarabokat "felszednie", amelyek lehetővé teszik, hogy később visszajusson az organellumba, és nem utolsósorban mindezenközben működőképesnek maradnia. Minden valószínűség szerint nem is sikerült általában elsőre az "átugrás", temérdek sikertelen kísérlet kellett minden egyes sikeres endoszimbiotikus géntranszferhez. Ami viszont fel is veti a kérdést, hogy a sikertelen alkalmakat is beleszámítva, mennyire gyakori a plasztisz, ill. mitokondriális eredetű gének átjutása a sejtmagba?
Ennek eldöntésére egy szellemes kísérletet végzett párhuzamosan több csoport is pár évvel ezelőtt. A kísérleti körülmények apró részletei eltértek, de a lényeg minden esetben kb. így foglalható össze: olyan transzgénikus növényeket hoztak létre, amelyek kloroplasztiszának a genomjába építenek be két antibiotikum rezisztencia gént (neo és aadA, hogy miként hatnak, az most mindegy). Az egyiknek (aadA) prokarióta-típusú promotere van, így a plasztiszban át tud íródni, mert az organellum baktériumokra hajazó transzkripció apparátusa felismeri. A másik (neo) azonban egy eukarióta promoterrel van ellátva, így csendben marad. (Mindez az alábbi ábra (a.) paneljén van összefoglalva.)
Ahhoz, hogy a neo meg tudjon nyilvánulni, az egész gén át kell kerüljön a sejtmagba, ahol aztán az eukarióta promoter már működőképes lesz (b.). Ha tehát azt nézzük, milyen valószínűséggel jelennek meg neo rezisztens növények, akkor népi képet kaphatunk arról, hogy mennyire gyakori az endoszimbiotikus géntranszfer egy élő növényben.
Az eredmények szerint szomatikus sejtekben (levélben) 1 : 5 millió volt az átugrás gyakorisága, míg pollen sejtekben 1 : 16,000. (A különbség egyik oka az lehet, hogy pollenképződés közben lebomlik a plasztisz és a felszabaduló DNS nagyobb eséllyel juthat a sejtmagba, így ott megnő a géntranszfer esélye is.) Figyelembe véve, hogy ez egy nyilvánvaló (nagyságredbeli) alábecslése az összes géntranszfer eseménynek - hiszen azokat az eseteket, amikor nem a teljes rezisztencia gén, vagy a plasztisz genom egy teljesen más szakasza került át, nem "látjuk" -, nem túlzás kijelenteni, hogy minden egyes növényben több ilyen kontrollálatlan géntranszfer játszódik le, különös tekintettel a hímivarsejtekre.
![]()
Érdekes módon a transzfer eredménye nem túl stabil, gyakran egyetlen generáció után elvesződik a rezisztencia gén (ezt önbeporzó növényeken mérték ki). Mivel, mint azt előbb részleteztem, sok mindennek kell egyszerre bekövetkeznie, hogy egy funkcionális géntranszfer bekövetkezzen, ez persze nem sokkolóan meglepő.
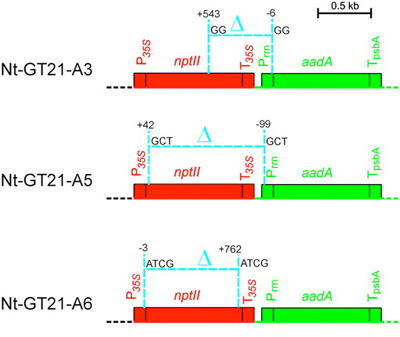 De a meglevő kísérleti rendszert tovább is lehet használni: azt is vizsgálhatjuk, hogy milyen körülmények eredményezhetnek egy átíródó plasztisz-eredetű gént. Ehhez előbb "kikeresztezéssel" olyan vonalakat hoztak létre, amelyek csak a sejtmagjukban tartalmazzák a transzgénikus kazettát, a plasztisz-genomjukban nem (c.). Majd azt nézték, hogy mikor jelentkezik a spectinomyicin rezisztencia - amihez az alapból prokarióta promotert hordozó aadA valahogy szert kell tegyen egy eukarióta jellegű promoterre. Szinte minden dokumentált eset azzal járt, hogy a transzgénikus kazettából az előtte levő rezisztencia gén (a mellékelt ábrán nptIII, fent (d.) pedig neo) egy deléció révén kitörlődött és a promótere így "átadódott" az aadA-nak.
De a meglevő kísérleti rendszert tovább is lehet használni: azt is vizsgálhatjuk, hogy milyen körülmények eredményezhetnek egy átíródó plasztisz-eredetű gént. Ehhez előbb "kikeresztezéssel" olyan vonalakat hoztak létre, amelyek csak a sejtmagjukban tartalmazzák a transzgénikus kazettát, a plasztisz-genomjukban nem (c.). Majd azt nézték, hogy mikor jelentkezik a spectinomyicin rezisztencia - amihez az alapból prokarióta promotert hordozó aadA valahogy szert kell tegyen egy eukarióta jellegű promoterre. Szinte minden dokumentált eset azzal járt, hogy a transzgénikus kazettából az előtte levő rezisztencia gén (a mellékelt ábrán nptIII, fent (d.) pedig neo) egy deléció révén kitörlődött és a promótere így "átadódott" az aadA-nak.
Tanulság túl sok nincs, max. annyi, hogy minden egyes növényben folyamatosan alattomos és felügyeletlen géntranszferek zajlanak le, egy kvázi-bakteriális forrásból. Ha ennek gondolatától kiráz a hideg, akkor jobb a salátától gyorsan megszabadulni, és holnaptól zsírt enni zsírral, szendigáborosan.
Annyit viszont remélem sikerült szemléltetni, hogy a Greenpeace egyik legfőbb érve, ami alapján csuklóból elutasítják a génmódosítás gondolatát, egyszerűen az emberek ismerethiányára apellál. Nem egy valós problémát fogalmaznak meg, egyszerűen az ismeretlennel ijesztgetnek. Márpedig az idegentől való ódzkodásra, felületes és hibás előítéletekre alapozni egy kampányt morálisan nehezen védhető álláspont. Ha nem egy személytelen technológia démonizálásáról lenne szó, hanem egy hús-vér népcsoportéról, persze még nyilvánvalóbb (és persze veszélyesebb) lenne az egész. De így sem szép.
(A Captain Lemon karikatúra a Chadlonius.com-ról származik.)
Maliga P (2003) Plant biology: Mobile plastid genes. Nature 422: 31-32.
Bock R, Timmis JN (2008) Reconstructing evolution: gene transfer from plastids to the nucleus. Bioessays 30(6): 556-66.
Stegemann S, Bock R (2006) Experimental reconstruction of functional gene transfer from the tobacco plastid genome to the nucleus. Plant Cell 18(11): 2869-78.
Timmis JN, Ayliffe MA, Huang CY, Martin W (2004) Endosymbiotic gene transfer: organelle genomes forge eukaryotic chromosomes. Nat Rev Genet 5(2): 123-35.
Stegemann S, Bock R (2009) Exchange of genetic material between cells in plant tissue grafts. Science 324: 649-51.